You might also like
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (890)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- A Passage To AfricaDocument2 pagesA Passage To AfricaXentrico123100% (1)
- EulogyDocument4 pagesEulogyDianne Yco100% (1)
- DM's Kit - On The Trail of TyrannyDocument50 pagesDM's Kit - On The Trail of Tyrannycengiz isyan100% (2)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- Prehistoric Compendium - GM BinderDocument95 pagesPrehistoric Compendium - GM BinderJacob Andersen-Lum0% (1)
- Auset-Ausar or Isis-Osiris PDFDocument5 pagesAuset-Ausar or Isis-Osiris PDFBrett Blanks100% (1)
- Legal Medicine 2018Document7 pagesLegal Medicine 2018german guazaNo ratings yet
- La1 La 2 PDFDocument8 pagesLa1 La 2 PDFChristian Rodriguez0% (1)
- Robert Porter Murder Incident ReportDocument2 pagesRobert Porter Murder Incident ReportThe Dallas Morning News100% (1)
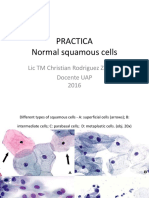
- Normal Squamous Cervical CellsDocument34 pagesNormal Squamous Cervical CellsChristian RodriguezNo ratings yet
- Citometria de FlujoDocument6 pagesCitometria de FlujoChristian RodriguezNo ratings yet
- Atlas Normal Squamous CellsDocument34 pagesAtlas Normal Squamous CellsChristian Rodriguez100% (1)
- Agua PDFDocument5 pagesAgua PDFChristian RodriguezNo ratings yet
- Paradise Lost - As I DieDocument5 pagesParadise Lost - As I DieWARLOCKNo ratings yet
- Crossing The Bar by Alfred Lord TennysonDocument7 pagesCrossing The Bar by Alfred Lord TennysonanushkahazraNo ratings yet
- Summary:: Keywords: Necropolis, XVIDocument4 pagesSummary:: Keywords: Necropolis, XVIIuliana GherbovetNo ratings yet
- The New Trauma Score (NTS) : A Modification of The Revised Trauma Score For Better Trauma Mortality PredictionDocument9 pagesThe New Trauma Score (NTS) : A Modification of The Revised Trauma Score For Better Trauma Mortality PredictionstellaNo ratings yet
- Paul Raj DeathDocument1 pagePaul Raj DeathJeya kumarNo ratings yet
- The City Stained Red by Sam Sykes ExtractDocument19 pagesThe City Stained Red by Sam Sykes ExtractOrion Publishing Group0% (1)
- Trifles EssayDocument3 pagesTrifles Essayapi-317562668No ratings yet
- Othello Final PaperDocument6 pagesOthello Final Paperapi-211233578No ratings yet
- Hrry Potter ND The WRTH of The FllenDocument619 pagesHrry Potter ND The WRTH of The FllenhheeaatthhNo ratings yet
- Format+for+enrollment+of+Sponsorship On 13.03.2023Document3 pagesFormat+for+enrollment+of+Sponsorship On 13.03.2023manichandu pNo ratings yet
- Death mask detailsDocument4 pagesDeath mask detailstanitartNo ratings yet
- Matt. Odietus Interview 2Document3 pagesMatt. Odietus Interview 2skalpsoloNo ratings yet
- What Are The Standard Puzzles Asked in Programming Interviews - QuoraDocument6 pagesWhat Are The Standard Puzzles Asked in Programming Interviews - QuoraRahul BhayaNo ratings yet
- PoemsDocument5 pagesPoemsapi-342778315No ratings yet
- Book of The DeadDocument13 pagesBook of The DeadCatherine TheGreatNo ratings yet
- Death Penalty Case NegDocument67 pagesDeath Penalty Case NegIanNo ratings yet
- Cause of Death Rate/ 10,000 Pop (TB) 2. Swaroop's IndexDocument2 pagesCause of Death Rate/ 10,000 Pop (TB) 2. Swaroop's IndexLeslie CruzNo ratings yet
- Alaban Vs CADocument14 pagesAlaban Vs CABerNo ratings yet
- The Tiger KingDocument3 pagesThe Tiger Kinghrithik67% (3)
- Montaer Vs Sharia District CourtDocument5 pagesMontaer Vs Sharia District CourtLloyd SanchezNo ratings yet
- False Starts and Second GuessesDocument169 pagesFalse Starts and Second Guessesyotoob100% (4)
- Who was Pedro Alonso Lopez, the serial killer known as "The Monster of the AndesDocument5 pagesWho was Pedro Alonso Lopez, the serial killer known as "The Monster of the Andesdonny waltNo ratings yet
- Child Sacrifice ReportDocument96 pagesChild Sacrifice ReportprowlingNo ratings yet