Professional Documents
Culture Documents
Review: Conceptual Framework, Defi Nitions, and Animal Models
Uploaded by
MarinaOriginal Title
Copyright
Available Formats
Share this document
Did you find this document useful?
Is this content inappropriate?
Report this DocumentCopyright:
Available Formats
Review: Conceptual Framework, Defi Nitions, and Animal Models
Uploaded by
MarinaCopyright:
Available Formats
Review
Neurobiology of addiction: a neurocircuitry analysis
George F Koob, Nora D Volkow
Lancet Psychiatry 2016; Drug addiction represents a dramatic dysregulation of motivational circuits that is caused by a combination of
3: 76073 exaggerated incentive salience and habit formation, reward decits and stress surfeits, and compromised executive
National Institute on Alcohol function in three stages. The rewarding eects of drugs of abuse, development of incentive salience, and development
Abuse and Alcoholism
of drug-seeking habits in the binge/intoxication stage involve changes in dopamine and opioid peptides in the basal
(G F Koob PhD) and National
Institute on Drug Abuse ganglia. The increases in negative emotional states and dysphoric and stress-like responses in the withdrawal/
(N D Volkow MD), National negative aect stage involve decreases in the function of the dopamine component of the reward system and
Institutes of Health, Rockville, recruitment of brain stress neurotransmitters, such as corticotropin-releasing factor and dynorphin, in the
MD, USA
neurocircuitry of the extended amygdala. The craving and decits in executive function in the so-called preoccupation/
Correspondence to:
anticipation stage involve the dysregulation of key aerent projections from the prefrontal cortex and insula, including
Dr George F Koob, National
Institute on Alcohol Abuse and glutamate, to the basal ganglia and extended amygdala. Molecular genetic studies have identied transduction and
Alcoholism, 5635 Fishers Lane, transcription factors that act in neurocircuitry associated with the development and maintenance of addiction that
Room 2001, Suite 2000, might mediate initial vulnerability, maintenance, and relapse associated with addiction.
Rockville, MD 20852, USA
gkoob@scripps.edu
Conceptual framework, denitions, and animal In 2013, DSM-51 combined what was previously
models conceptualised as two separate and hierarchical disorders
Drug addiction can be dened as a chronically relapsing (substance abuse and substance dependence) into one
disorder, characterised by compulsion to seek and take construct, dening substance use disorders on a range
the drug, loss of control in limiting intake, and from mild to moderate to severe, with the severity of an
emergence of a negative emotional state (eg, dysphoria, addiction depending on how many of the established
anxiety, irritability) when access to the drug is prevented. criteria apply.
From a diagnostic perspective, the term addiction is now Although much of the early research with animal
encompassed by the term substance use disorders. models focused on the acute rewarding eects of drugs of
abuse, focus is shifting to the study of chronic drug
administration-induced brain changes that decrease the
Key messages
threshold for relapse, which corresponds more closely to
Substance use disorders are complex, multistage diseases that are characterised by human imaging studies of individuals who have substance
disturbances in three major neurocircuits: (i) basal ganglia-driven binge/intoxication use disorders. One of the main goals of neurobiological
stage, (ii) extended amygdala-driven withdrawal/negative aect stage, and (iii) research is to understand changes at the molecular,
prefrontal cortex-driven preoccupation/anticipation stage. cellular, and neurocircuitry levels that mediate the
In these three domains, neurotransmitter-specic and neuromodulator-specic transition from occasional, controlled substance use to
neuroplastic changes are seen in 18 subsystems, including the ascending loss of control in drug intake and chronic addiction.2
mesocorticolimbic dopamine system, corticotropin-releasing factor in the central Because only some substance users make this transition,
nucleus of the amygdala, and corticostriatal glutamate projections. neurobiological factors that inuence the diverse
During the binge/intoxication stage, previously neutral stimuli become associated individual dierences in drug responses have also
with drug availability, thereby gaining incentive salience and promoting habit attracted increasing interest. Cogent arguments have been
formation that fosters excessive drug seeking via increases in dopamine and made that addictions are similar to other chronic relapsing
glutamate neurotransmission. disorderswith individual dierences in responses to the
The binge/intoxication stage triggers opponent-process responses that diminish same exogenous challenges and limited ecacy of
reward function via dopamine and opioid peptide decits and increased brain stress treatmentsuch as diabetes, asthma, and hypertension.3
system activity through the engagement of corticotropin-releasing factor and The purpose of this Review is twofold. First, we aim to
dynorphin. elaborate a heuristic framework based on the neuro-
Excessive drug intake also drives parallel decits in executive function via the psychopharmacological and brain imaging phenotype
dysregulation of glutamatergic, GABAergic, and dopaminergic neuronal networks in of addiction in the context of three functional domains
the prefrontal cortex, which perpetuate the dysregulation of reward and stress (incentive salience, negative emotional states, and
function and induce compulsive drug use. executive function) mediated by three major neuro-
During the preoccupation/anticipation stage, heightened drug cue-induced incentive biological circuits (basal ganglia, extended amygdala,
salience can act against a background of low reward system function and high stress and prefrontal cortex). Second, we aim to identify
system function, engaging a powerful combination of positive and negative neurochemically dened mini circuits that can
reinforcement processes that drive pathological drug seeking. independently or interactively mediate functional
Molecular genetic mediation and epigenetic changes in these same circuits provide neuroplasticity within the three major circuits to
pre-existing vulnerabilities to addiction, increased susceptibility to environmental risk produce incentive salience and compulsive-like habits,
factors, and targets for development of novel treatments and resilience to relapse. negative emotional states of low reward and excessive
stress, and compromised executive function. Building
760 www.thelancet.com/psychiatry Vol 3 August 2016
Review
on previous identication of the three overall domains, use is initiated in adolescence, and the distinct drug use
this Review provides a framework for integration of the trajectories that are observed in men and women. The
ever-expanding neuroplastic complexity of motivational neurobiological mechanisms involved in the stages of the
systems that are involved in addiction and for addiction cycle can be conceptualised as domains, with a
identication of new targets for diagnosis, treatment, focus on specic brain circuits, the molecular and
and prevention of addiction. neurochemical changes in those circuits during the
Addiction can be conceptualised as a three-stage, transition from drug taking to addiction, and the way in
recurring cyclebinge/intoxication, withdrawal/negative which those changes persist in the vulnerability to relapse.12
aect, and preoccupation/anticipation (craving)that Equally convincing, animal models of the specic stages of
worsens over time and involves neuroplastic changes in the addiction cycle can be paralleled by human laboratory
the brain reward, stress, and executive function systems.46 models13 and studied with neuroimaging.14
Derived from a conuence of information from social
psychology of human self-regulation failure, psychiatry, Neurobiological mechanisms of the binge/
and brain imaging, these three stages provide a heuristic intoxication stage
framework for the study of the neurobiology of addiction.4,5 Drug reward
A denition of impulsivity is a predisposition toward Drugs of abuse activate brain reward systems, and
rapid, unplanned reactions to internal and external research on drug addiction has in large part dened the
stimuli without regard for the negative consequences of neurocircuitry of reward. This line of investigation is
these reactions to themselves or others.7 A denition of fundamental because changes to how the drug-induced
compulsivity is the manifestation of perseverative, reward system is activated are key to the understanding
repetitive actions that are excessive and inappropriate.8 of the development of addiction.4 Reward is dened
Impulsive behaviours are often accompanied by feelings herein as any event that increases the probability of a
of pleasure or gratication, but compulsions in disorders
such as obsessive-compulsive disorder are often
performed to reduce tension or anxiety from obsessive
thoughts.8 In this context, individuals move from
impulsivity to compulsivity, and the drive for drug-taking
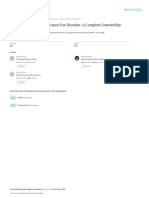
behaviour is paralleled by shifts from positive to negative PFC
(ACC,
Dorsal striatum
Motor cortex
reinforcement (gure 1). However, impulsivity and inferior
PFC, 1
3 4
2
Withdrawal/
Binge/intoxication lateral
compulsivity can coexist, and frequently do so in the OFC) NAc-VTA negative aect
la
12 13
nu
5
dierent stages of the addiction cycle.8
be
8 9
Ha
E 7
Understanding of the neurobiology of addiction has PFC amxten 17
(medial OFC) yg ded 18
da
la
progressed through the study of animal models10 and, 16 14
15
6 10 11
more recently, through brain imaging studies in Insula
individuals with addiction. Although no animal model of
addiction fully emulates the human condition, they permit
Incentive Reward decit
investigations of specic signs or symptoms that are and stress surfeit
salience
associated with the psychopathological condition. If the
model adequately mimics the phenomenology observed in
humans as they transition from experimentation to Preoccupation/
addiction, then it is more likely to have construct or anticipation
predictive validity.11 The phenomena under study can be
models of dierent systems (genetic, epigenetic, trans- Executive function
cription, cellular, and network), psychological constructs
(positive and negative reinforcement), symptoms outlined
by psychiatric nosology (craving, hypohedonia, and Neurocircuits Synaptic Molecules
systems
dysphoria), and stages of the addiction cycle.4 Recently
Neuroadaptation
developed animal models take advantage of individual and
strain diversity in responses to drugs, incorporate complex
environments with access to and choices of alternative Figure 1: Model of interacting circuits in which disruptions contribute to compulsive-like behaviours
underlying drug addiction
reinforcers, and test eects of stressful stimuli, allowing The overall neurocircuitry domains correspond to three functional domains: binge/intoxication (reward and
the investigation of neurobiological processes that underlie incentive salience: basal ganglia [blue]), withdrawal/negative aect (negative emotional states and stress:
the risk for addiction and environmental factors that extended amygdala and habenula [red]), and preoccupation/anticipation (craving, impulsivity, and executive
provide resilience against vulnerability. Animal models function: PFC, insula, and allocortex [green]). Arrows depict major circuit connections between domains, and
numbers refer to neurochemical and neurocircuit-specic pathways known to support brain changes that
have also started to explore the inuence of developmental contribute to the allostatic state of addiction. PFC=prefrontal cortex. ACC=anterior cingulate cortex.
stage and sex in drug response to better understand the OFC=orbitofrontal cortex. NAc-VTA=nucleus accumbens-ventral tegmental area. Modied from Koob and Volkow
greater vulnerability to substance use disorders when drug (2010)9 with permission from Nature Publishing Group.
www.thelancet.com/psychiatry Vol 3 August 2016 761
Review
Response Neurotransmitter
Binge/intoxication Binge/intoxication
Dopamine15 Increase Ventral tegmental area (circuit 1) 43 Glutamate
Opioid peptides 16
Increase Ventral tegmental area (circuit 2) 44 -aminobutyric acid
Serotonin17 Increase Dorsal striatum (circuit 3) 45 Dopamine
-aminobutyric acid18 Increase Dorsal striatum (circuit 4) 46 Glutamate
Acetylcholine19 Increase Withdrawal/negative aect
Withdrawal/negative aect Ventral tegmental area (circuit 5) 47 Corticotropin-releasing factor
Corticotropin-releasing factor20
Increase Central nucleus of amygdala (circuit 6) 20 Corticotropin-releasing factor
Dynorphin21 Increase BNST (circuit 7) 22 Norepinephrine
Norepinephrine22 Increase Nucleus accumbens shell (circuit 8) 21 Dynorphin
Hypocretin (orexin) 23 Increase Habenula (circuit 9) 48 Acetylcholine
Substance P24 Increase Central nucleus of amygdala (circuit 10) 49 Neuropeptide Y
Dopamine25 Decrease Central nucleus of amygdala (circuit 11) 29 Endocannabinoids
Serotonin 17
Decrease Preoccupation/anticipation
Opioid peptide receptors26 Decrease Prefrontal cortex (circuit 12) 32 Glutamate
Neuropeptide Y 27 Decrease Prefrontal cortex (circuit 13) 50 -aminobutyric acid
Nociceptin28 Decrease Hippocampus (circuit 14) 51 Glutamate
Endocannabinoids29 Decrease Basolateral amygdala (circuit 15) 52 Glutamate
Oxytocin30 Decrease BNST (circuit 16) 53 Corticotropin-releasing factor
Preoccupation/anticipation BNST (circuit 17) 53 Norepinephrine
Dopamine31 Increase Insula (circuit 18) 54 Corticotropin-releasing factor
Glutamate32 Increase
BNST=bed nucleus of the stria terminalis.
Hypocretin (orexin) 23 Increase
Serotonin 17
Increase Table 2: Molecular neurocircuits as focal points for neuroplasticity in
addiction
Corticotropin-releasing factor33 Increase
Table 1: Neurotransmitter systems involved in the neurocircuitry of
addiction stages and functional domains
has the understanding of the relevant neurotransmitters
and neuromodulators, which include not only dopamine
response with a positive hedonic component. A and opioid peptides but also -aminobutyric acid (GABA),
principal focus of research on the neurobiology of the glutamate, serotonin, acetylcholine, and endocannabinoid
rewarding eects of abused drugs has been the origins systems that act at the level of either the ventral tegmental
and terminal areas of the ascending mesocorticostriatal area or nucleus accumbens (gure 1; tables 1, 2 for
dopamine systems that have a key role in the rewarding neurotransmitter systems and specic neurocircuits
properties of nearly all drugs of abuse (table 1).34 In involved). Balanced circuits result in proper inhibitory
humans, positron emission tomography studies have control and decision making and normal functioning of
shown that intoxicating doses of alcohol and drugs reward, motivation, stress, and memory circuits. These
release dopamine and opioid peptides into the ventral circuits also interact with circuits that are involved in
striatum,16,35 and that fast and steep release of dopamine mood regulation, including stress reactivity (which
is associated with the subjective sensation of the so- involves the amygdala, hypothalamus, and habenula) and
called high.36 This is because fast and steep increases in interoception (which involves the insula and anterior
dopamine activate low-anity dopamine D1 receptors, cingulate cortex and contributes to the awareness of
which are necessary for the rewarding eects of drugs37 negative emotional states). Drugs of abuse usurp
and for triggering conditioned responses.38 By contrast, executive function circuits, motivational circuits, and
dopamine stimulation of high-anity dopamine D2 stress circuits via multiple neurotransmitter-specic
receptors is not sucient for drug reward,39,40 and these neuroplasticity circuits (tables 1, 2). Key neurotransmitters
receptors might even limit drug reward.41 In this that are implicated in these neuroadaptations include
respect, drugs emulate the increases in dopamine dopamine, enkephalins, glutamate, -aminobutyric acid,
triggered by phasic dopamine ring, which are the norepinephrine, corticotropin-releasing factor (CRF),
ring frequencies of dopamine neurons associated dynorphin, neuropeptide Y, and endocannabinoids
with rewarding stimuli.42 (tables 1, 2).
The specic circuitry that is associated with drug reward
has been broadened to include many neural inputs and Incentive salience
outputs that interact with the basal forebrain. As the Drugs of abuse have a profound eect on the response to
understanding of the relevant circuits has evolved, so too previously neutral stimuli to which the drugs become
762 www.thelancet.com/psychiatry Vol 3 August 2016
Review
paired. This phenomenon, called conditioned reinforce- within the incentive salience process appear to drive
ment, can be dened as when a previously neutral stimulus dopamine signalling to maintain motivation to take the
reinforces or strengthens behaviours through its drug, even when its pharmacological eects lessen.57,60,61 By
association with a primary reinforcer and becomes a contrast, parallel impairments in executive function that
reinforcer in its own right. Such cues can be contextual are mediated by the prefrontal cortex might be linked to
and predictive, and the process of conditioned changes in tonic dopamine cell ring that result in lower
reinforcement entails not only approach to salient cues but but more stable dopamine levels in the dopamine
also instrumental responding to turn on the cues, in view projections to the prefrontal cortex.62,63 This mechanism
of their own rewarding (conditioned reinforcing) implicates lower-anity D1 receptors, which stimulate
properties. As such, conditioned reinforcement as a cyclic adenosine monophosphate (cAMP) signalling, as
construct preceded and laid the foundation for incentive being involved in both acute drug reward and conditioning,
salience.55 Incentive salience can be dened as motivation because both induce spikes in dopamine release. By
for rewards derived from both ones physiological state and contrast, D2 receptors, which inhibit cAMP signalling, are
previously learned associations about a reward cue that is stimulated by both phasic and tonic dopamine and are not
mediated by the mesocorticolimbic dopamine system.56 deemed necessary for drug reward.40,64 D3 receptors, which
Both conditioned reinforcement and incentive salience are high-anity dopamine receptors and co-localise in the
provide constructs for what underlies cue-induced drug nucleus accumbens with D1 receptors, are also associated
seeking, self-administration behaviours, and, conceivably, with drug-seeking behaviour.65
the transition to habit-like compulsive drug seeking.
The neurobiology that underpins these processes has Neurobiological mechanisms of the withdrawal/
received much research interest. One particularly negative aect stage
inuential series of studies in non-human primates The withdrawal/negative aect stage consists of key
indicated that midbrain dopamine cells initially red in motivational elements, such as chronic irritability,
response to a novel reward. After repeated exposure, the emotional pain, malaise, dysphoria, alexithymia, states of
neurons stopped ring during predictable reward stress, and loss of motivation for natural rewards. Across
delivery and instead red when they were exposed to all major drugs of abuse, this stage is characterised in
stimuli that were predictive of the reward.57 A new laboratory animals by elevations in reward thresholds (ie,
rewardand subsequently the cues associated with the decreased reward) during withdrawal. In animal models
rewardtriggered phasic dopamine cell ring and of the transition to addiction, elevations in brain reward
activation of dopamine D1 receptors, which are necessary thresholds occur that temporally precede and are highly
for conditioning to occur (table 2, circuits 14).58 This correlated with escalation in drug intake.20 During acute
process allows previously neutral stimuli to become and protracted withdrawal from chronic administration
endowed with incentive salience and strengthens the of all drugs of abuse, increases in stress and anxiety-like
learned association with repeated exposure to the cues, responses also occur that contribute greatly to the
which creates strong motivation to seek a reward. malaise of abstinence and protracted abstinence
Phasic dopamine signalling induced by drug (table 1).66 Human brain imaging studies have reported
administration can also trigger neuroadaptations in basal decreases in the sensitivity of brain reward circuits to
ganglia circuits. Here, drug-induced phasic dopamine stimulation by natural rewards during the withdrawal/
release triggers the ability of drug-paired cues to increase negative aect stage.67
dopamine levels. Activation of the ventral striatum Chronic drug exposure-induced neurochemical
leads to the recruitment of striatal-globus pallidal- changes in systems that are implicated in acute drug
thalamocortical loops that engage the dorsal striatum, reward are called within-system neuroadaptations.
resulting in habit formation45 and triggering what is Within-system neuroadaptations can be dened as those
hypothesised to underlie compulsive responding for in which the primary cellular response element to the
drugs (table 2, circuits 3 and 4).59 Key synaptic changes drugadapt[s] to neutralize the drugs eects; persistence
involve glutamate-modulated N-methyl-D-aspartate of the opposing eects after the drug disappears
(NMDA) receptors and -amino-3-hydroxy-5-methyl-4- produce[s] the withdrawal response.68 Such changes
isoxazolepropionic acid (AMPA) receptors in gluta- include decreases in dopaminergic and serotonergic
matergic projections from the prefrontal cortex and transmission in the nucleus accumbens during drug
amygdala to the ventral tegmental area and nucleus withdrawal as measured by in-vivo microdialysis in rats69
accumbens (table 2, circuit 4).32,43,46 and brain imaging studies in humans, in which
The ability of conditioned cues to recruit these circuits amphetamine-induced or methylphenidate-induced
augments progression through the addiction cycle and striatal dopamine responses are 50% lower in detoxied
helps explain intense desire for the drug (ie, craving) and abusers and 80% lower in active abusers, and
compulsive use when individuals with addiction are accompanied by lower self-reports of the drugs rewarding
exposed to drug cues or stressful environments that are eects relative to non-drug-abusing controls.25,35,61,70 Other
linked with negative emotional states. Conditioned cues observed changes include increases in opioid receptor
www.thelancet.com/psychiatry Vol 3 August 2016 763
Review
responsivity during opioid withdrawal,71,72 and decreases an expected reward.80,81 This hypothesis is also consistent
in GABAergic and increases in NMDA glutamatergic with the nding that 5 nicotinic acetylcholine receptors
transmission in the nucleus accumbens.73,74 Dierential in the habenula appear to modulate aversive responses to
regional changes in nicotinic receptor function in the large doses of nicotine48 and, along with habenular
nucleus accumbens and ventral tegmental area in 2 receptors, nicotine withdrawal (tables 1, 2, circuit 9).82
nicotine, alcohol, and other addictions have also been Anti-reward circuits are engaged as neuroadaptations
reported, implicating 42 nicotinic receptor subtypes.19,44 during the development of addiction, producing aversive
Such decreases in reward system function might persist or stress-like states. These aversive states are manifest
in the form of long-term biochemical changes that when the drug is removed during acute withdrawal but
contribute to the clinical syndrome of acute withdrawal also during protracted abstinence.2 Thus, the within-
and protracted abstinence and could also explain the loss system and between-system construct could be equally
of interest in normal, non-drug rewards (ie, narrowing of valid for the preoccupation/anticipation stage. The
the behavioural repertoire toward drugs and drug-related combination of decreases in reward function and
stimuli; tables 1, 2, circuit 5). increases in stress function in the motivational circuits
The emotional dysregulation that is associated with of the ventral striatum, extended amygdala, and habenula
the withdrawal/negative aect stage also involves a is a powerful trigger of negative reinforcement that
between-system neuroadaptation, in which neuro- contributes to compulsive drug-seeking behaviour and
chemical systems other than those involved in the addiction. Decreases in reward can be driven by
positive rewarding eects of drugs of abuse are recruited overactivation of the habenula or overactivation of the
or dysregulated by chronic activation of the reward dynorphin system in the ventral striatum, both of which
system.68 Both the hypothalamic-pituitary-adrenal axis can decrease dopamine neuron ring. Increases in
and brain stress system mediated by CRF are stress-like states and increased responsivity to stressors
dysregulated by the chronic administration of all major could be driven by the recruitment of CRF in the
drugs with dependence or abuse potential, with a amygdala and other extrahypothalamic stress systems.
common response of elevated adrenocorticotropic Compulsive-like habits might also derive from the so-
hormone, corticosterone, and amygdala CRF during called dark side of the withdrawal/negative aect stage.
acute withdrawal.20,75 As tolerance and withdrawal In a study of patients with obsessive-compulsive disorder,
develop, brain stress systems, such as CRF, patients showed enhanced avoidance habits compared
norepinephrine, and dynorphin, are recruited in the with controls following overtraining on a shock avoidance
extended amygdala and contribute to the development of task, providing support for a habit account of obsessive-
negative emotional states in withdrawal and protracted compulsive disorder.83 Whether a decient inhibitory
abstinence (tables 1, 2, circuits 68).20,22 As a result, the process in prefrontal regions or overactive goal-directed
concept of anti-reward was developed,2 based on the actions and habit processes in the basal ganglia mediate
proposal that opponent processes that are a general this eect remains to be determined, but the
feature of biological systems also act to limit reward.4 enhancement of avoidance habits provides another basis
Multiple circuits are likely to contribute to the for the contribution of activation of the stress axis to
hypothesised opponent-like processes. During acute compulsive-like responding.
withdrawal from all drugs of abuse, CRF increases in the Endogenous anti-stress systems also appear to buer
extended amygdala (tables 1, 2, circuit 6). Importantly, the brain stress systems and inuence vulnerability to
CRF receptor antagonists block both the anxiety-like, the development and perpetuation of addiction. Key
stress-like eects of drug withdrawal and excessive drug neurotransmitters that act in opposition to brain stress
taking during compulsive drug seeking in animals. systems include neuropeptide Y, nociceptin, and
Equally compelling, opioid receptor antagonists, when endocannabinoids (tables 1, 2, circuits 10 and 11).20,28,29,49,84
injected into the nucleus accumbens shell, can block the In particular, with chronic drug exposure, neuro-
development of compulsive drug seeking (tables 1, 2, adaptations in the endocannabinoid system, which
circuit 8).7678 Decreases in the release of dopamine in the modulates the response of the brain to stress,85 might
nucleus accumbens can also be driven by increases in contribute to the enhanced stress reactivity in addiction.
the activity of the dynorphin- opioid receptor system in Indeed, human brain imaging studies have reported
the ventral striatum and possibly increases in the activity reductions of cannabinoid CB1 receptors in cannabis
of CRF in the ventral tegmental area, which contributes abusers and alcoholics.86,87 Thus, the development of
to the negative emotional state associated with enduring aversive emotional states,88 mediated by
withdrawal and protracted abstinence.20,21 overactivation of the stress and anti-reward systems or
The lateral habenula plays a key role in mediating and underactivation of the anti-stress systems, might
encoding aversive states.79 Numerous studies indicate contribute to the crucial problem in drug addiction of
that the habenula is one of the regions that control the chronic relapse, in which individuals with addiction
decreases in dopamine neuron ring in the ventral return to compulsive drug taking long after acute
tegmental area that are associated with failure to receive withdrawal.2
764 www.thelancet.com/psychiatry Vol 3 August 2016
Review
Neurobiological mechanisms of the regulate incentive salience and conditioned behaviour
preoccupation/anticipation stage when a salient cue is presented to the individual.92,93
The preoccupation/anticipation stage has long been Evidence from rodent studies suggests that drug-induced
hypothesised to be a key element of relapse in humans, reinstatement is mediated by the circuit that links the
and denes addiction as a chronic relapsing disorder. prelimbic prefrontal cortex to the ventral striatum94 (for
Although this stage has often been linked to the construct correspondence with humans, see gure 2; tables 1, 2,
of craving, craving in itself has been dicult to measure circuits 12 and possibly 13). In rats, neurotransmitter
in human clinical studies and does not always correlate systems that are involved in drug-induced reinstatement
with relapse.89 Nevertheless, the stage in which the involve a glutamatergic projection from the prelimbic
individual reinstates drug-seeking behaviour after prefrontal cortex to the nucleus accumbens that is
abstinence remains a focus for identifying neurobiological modulated by dopamine activity through D1 and D2
mechanisms of relapse and the development of receptors in the frontal cortex. Cue-induced reinstatement
medications for treatment. also involves a glutamatergic projection from the prelimbic
Executive control over incentive salience is essential to prefrontal cortex, basolateral amygdala, and ventral
maintain goal-directed behaviour and the exibility of subiculum to the nucleus accumbens, and dopamine
stimulusresponse associations. In rats, the prefrontal modulation in the basolateral amygdala and dorsal
cortex (mainly prelimbic cortex, and some infralimbic striatum (tables 1, 2, circuits 14 and 15).51,52,95 By contrast, the
cortex) sends glutamatergic projections directly to stress-induced reinstatement of drug-related responding
mesocortical dopamine neurons in the ventral tegmental in animal models appears to depend on the activation of
area, thus exerting excitatory control over dopamine cell both CRF and norepinephrine in elements of the extended
ring and dopamine release in the prefrontal cortex90 (for amygdala (ie, central nucleus of the amygdala and bed
correspondence with humans, see gure 2). Given that nucleus of the stria terminalis; tables 1, 2, circuits 16
ventral tegmental area dopamine cells project heavily to and 17)22,33,53,9698 and the ventral tegmental area (tables 1, 2,
the basal ganglia, this frontal cortex glutamatergic circuit 9). Protracted abstinence, largely described in
projection might contribute to the development of alcohol dependence models, appears to involve overactive
incentive salience. Glutamatergic projections from the glutamatergic and CRF systems.99,100 Based on animal
prefrontal cortex to the caudate and ventral striatum also studies using rodents, increases in activity of the prefrontal-
modulate the control of the striatal-pallidal-thalamo- glutamatergic system could be hypothesised to elicit a
cortical system through both direct (D1 receptor- strong glutamatergic response that mediates craving-like
mediated) and indirect (D2 receptor-mediated) pathways. responses during the preoccupation/anticipation stage
Thus, the prefrontal cortex is in a good position to (tables 1, 2, circuits 10 and 11).
Rat Human
ACC Stimulus value ACC
Action value/cost Thalamus
dIPFC
Anticipation/availability DS GP
Context vIPFC
PL
Action inhibition vmPFC NAc
Emotion control BN HPC External context
ST
Outcome valuation CeA Insula
OFC Internal context
IL Drug subjective value
INS
OFC
Aective state
Stress
Incentive to action
Figure 2: Correspondence between rat and human brain regions relevant to the addiction process
Rats are commonly studied to unveil the neurobiological mechanisms of addiction because they have a well characterised central nervous system whose
neurochemical and molecular pathways in subcortical areas correspond reasonably well to those in humans. ACC=anterior cingulate cortex. PL=prelimbic cortex.
IL=infralimbic cortex. OFC=orbitofrontal cortex. INS=insula. dlPFC=dorsolateral prefrontal cortex. vlPFC=ventrolateral prefrontal cortex. vmPFC=ventromedial
prefrontal cortex. DS=dorsal striatum. GP=globus pallidus. NAc=nucleus accumbens. BNST=bed nucleus of the stria terminalis. CeA=central nucleus of the amygdala.
HPC=hippocampus. Modied with permission from George and colleagues (2012),50 Koob and colleagues (2014),91 and the National Academy of Sciences.
www.thelancet.com/psychiatry Vol 3 August 2016 765
Review
In humans, cue-induced craving appears to involve involves a widely distributed and more complex prefrontal
activation of similar circuits, in which cues that are cortexsubcortical circuitry. Positron emission tomography
associated with drugs and elements of non-drug studies have uncovered substantial reductions of
addictions produce activation of the prefrontal cortex, D2 receptor availability in the striatum in individuals with
including the dorsolateral prefrontal cortex, anterior addiction that persist for months after protracted
cingulate gyrus, and medial orbitofrontal cortex.101105 detoxication, which have also been observed in preclinical
Imaging studies have also revealed that cues associated studies in rodents and non-human primates with repeated
with cocaine craving increase dopamine release in the drug exposure.122 These low levels of striatal D2 receptors
striatum, amygdala, and prefrontal cortex (anterior have been associated with decreases in baseline glucose
cingulate and medial orbitofrontal cortex) and opioid metabolism (ie, a marker of brain function) in the
peptides in the anterior cingulate and frontal cortex.9,31,106,107 prefrontal cortex (including dorsolateral prefrontal cortex,
Indeed, imaging studies have revealed baseline decreases anterior cingulate gyrus, and medial orbitofrontal
in orbitofrontal (medial and lateral) and anterior cingulate cortex),35,123,124 impulsivity in methamphetamine abusers,125
(ventral and dorsal) function and dopamine function and compulsive cocaine administration in rodents.15
during dependence, but reactivation of dopamine and Consistent with this hypothesis, the converse is also
reward system function during acute craving episodes.108 possible. Methamphetamine-dependent individuals with
Concomitant with prefrontal activation of a craving striatal D2 receptor availability within the normal range
system that is mediated by glutamate, human imaging had better treatment outcomes, potentially reecting a
studies have reported decits in executive function that greater ability to acquire new reward-related behaviours
are reected by decreases in frontal cortex activity that and the pursuit of healthy goals.126
interfere with decision making, self-regulation, inhibitory Two opposing systems can therefore be postulated: a Go
control, and working memory,109 and might involve system and a Stop system. The Go system could drive
disrupted GABAergic activity in the prefrontal cortex craving and engage habits via the basal ganglia.105,127 For
(table 2, circuit 13).50 For example, alcoholics exhibit example, steeper delayed discounting and cocaine craving
impairments in the maintenance of spatial information, in cocaine dependence is associated with increased
disruption of decision making, and impairments in connectivity in the network linking the medial prefrontal
behavioural inhibition. Indeed, Volkow and colleagues36 cortex and anterior cingulate cortex with the ventral
and Goldstein and Volkow14 have described a syndrome striatum and in the network linking the insula with the
that is characterised by excessive salience to drug-paired dorsal striatum.128 Activation of the dorsal anterior
cues, decreases in the responsiveness to non-drug cingulate is also associated with fear conditioning in post-
rewards, and decreases in the ability to inhibit maladaptive traumatic stress disorder.129 The Stop system could control
behaviour. Such frontal cortex-derived executive function assessment of the incentive value of choices and
disorders in addiction have been linked to decits in the suppression of aective responses to negative emotional
ability of behavioural treatments to eect recovery.110 signals.130132 Under this framework, a Stop system would
Cravings for food, cocaine, and nicotine are also inhibit the Go craving system and stress system. For
related to middle insular function.111113 Tobacco smokers example, in post-traumatic stress disorder there is
with damage to the insula (but not control smokers with substantial evidence of hypoactivity in the ventromedial
extra non-insular lesions) were able to stop smoking prefrontal cortex and hyperactivity in the central nucleus
easily and without experiencing cravings or relapse.114 of the amygdala that reects an inhibitory connection
The insula appears to have an interoceptive function between the ventromedial prefrontal cortex and central
that integrates autonomic and visceral information with nucleus of the amygdala.133,134 Indeed, successful cognitive
emotion and motivation, thus providing conscious inhibition of craving in cocaine abusers has been
awareness of these urges.54 Brain lesion studies suggest associated with inhibition in the right medial orbitofrontal
that the ventromedial prefrontal cortex and insula are cortex and activation in the right inferior frontal cortex.135
necessary components of the distributed circuits that
support both emotional115 and moral116 decision making. Molecular and genetic treatment targets within
Consistent with this hypothesis, imaging studies have brain circuits associated with addiction
reported dierential activation of the insula during The neuroplastic changes outlined previously are
craving, possibly reecting interoceptive cues and triggered and sustained by molecular and cellular
hypothesised to involve CRF activation (table 2, adaptations that can presumably also interact with
circuit 18).54,117,118 Accordingly, the reactivity of this brain genetic and environmental vulnerability to addiction. In
region has been suggested to serve as a biomarker to the binge/intoxication stage, both signal transduction
help predict relapse.119 mechanisms and changes in gene transcription have
The capacity to inhibit prepotent responses is a major been identied. For example, chronic exposure to a wide
contributor to an individuals vulnerability to addiction variety of abused drugs upregulates cAMP formation,
because it modulates the ability to avoid inappropriate cAMP-dependent protein kinase A (PKA) activity, and
behaviours.120,121 Such a conceptual inhibition system PKA-dependent protein phosphorylation in the nucleus
766 www.thelancet.com/psychiatry Vol 3 August 2016
Review
accumbens. Numerous interventions that tonically The kinase mammalian (mechanistic) target of
activate nucleus accumbens cAMP/PKA signalling rapamycin complex 1 (mTORC1) belongs to the
promote escalations in drug self-administration or phosphoinositide 3-kinase (PI3K)-related kinase (PIKK)
compulsive-like drug-seeking behaviour, and the family and plays a key role in the dendritic translation of
upregulation of a postsynaptic Gs/cAMP/PKA signalling synaptic proteins. As such, mTORC1 plays a major role
pathway in the nucleus accumbens might constitute a in the molecular mechanisms associated with learning
critical neuroadaptation that is central to the and memory147 and is involved in several brain disorders,
establishment and maintenance of the addicted state.136,137 including epilepsy, Parkinsons disease, Alzheimers
These changes in signal transduction can trigger longer- disease, and addiction.148,149 Exposure to drug and alcohol
term molecular neuroadaptations via transcription factors cues activates the mTORC1 pathway in the hippocampus,
that modify gene expression. A well characterised example frontal cortex, nucleus accumbens, and amygdala.149 Even
is that chronic exposure to various drugs of abuse increases more compelling, blockade of the mTORC1 pathway
the activation of cAMP response element binding protein systemically blocked the reconsolidation of cocaine
in the nucleus accumbens and deactivates it in the central memories,150 and blockade of mTORC1 in the amygdala
nucleus of the amygdala. Introduction of cAMP response blocked the reconsolidation of alcohol memories;151 these
element binding protein in the nucleus accumbens results suggest a potential molecular target for the
decreases the reinforcing value of natural and drug treatment of relapse.
rewards, and this change plausibly contributes to Heritability of addictions is 4060%, much of which is
withdrawal/negative aect stage-related decreases in caused by genetic variations that aect underlying
reward pathway function, which leave a drug-abstinent neurobiological mechanisms, and thus is consistent with
individual in an amotivational, dysphoric, or depressed- there being common pathways to dierent addictions.152
like state.138,139 These substance-related changes in Genes have been identied that convey vulnerability at
susceptibility to negative emotional states might begin all three stages of the addiction cycle, and salient
early: alcohol use during adolescence might lead to candidates are discussed within this conceptual
epigenetic modications that alter amygdalar gene framework. However, a more comprehensive analysis is
expression and dendritic density, increasing susceptibility beyond the scope of this Review. In the binge/intoxication
to anxiety-like behaviours and alcohol ingestion in stage, several genes have been identied in animals as
adulthood.140 key to drug responses, and their modications strongly
Substance-induced changes in transcription factors aect drug self-administration.37,153155 Notably, in animal
can also produce competing eects on reward function.141 models, dopamine D1 receptor knockout rats will not self-
For example, repeated substance use activates administer cocaine37 and opioid receptor knockout
accumulating levels of FosB, and animals with elevated mice will not show the rewarding eects of opioids.153 In
FosB exhibit exaggerated sensitivity to the rewarding humans, only a few specic genes have been identied
eects of drugs of abuse, leading to the hypothesis that with polymorphisms (alleles) that either predispose an
FosB might be a sustained molecular trigger or switch individual to or protect an individual from drug
that helps initiate and maintain a state of addiction.141,142 addiction,156 but the number is growing. For example,
Next-generation sequencing methods have shown that genome-wide association studies have implicated two
repeated cocaine exposure leads to histone modications acetylcholine receptors, the 4 nicotinic receptor
that act in a combinational fashion to create chromatin subunit157 and 5 nicotinic receptor subunit,158 in the
signatures that strikingly alter pre-mRNA splicing, an vulnerability to nicotine dependence and a single-
eect that is necessary for the expression of cocaine- nucleotide polymorphism associated with regulating the
induced conditioned place preference.143 tracking and gating properties of AMPA-selective
The heightened risk of cue-induced cocaine seeking glutamate receptors (CNIH3) in opioid dependence.159
(incubation of craving) that occurs during prolonged Some of the polymorphisms associated with
withdrawal (protracted abstinence in humans) has vulnerability for the binge/intoxication stage interfere
provided new insights into the molecular basis of with drug metabolism. For example, specic alleles for
vulnerability to relapse. During incubation in animal genes that encode alcohol dehydrogenase (ADH1B) and
models, AMPA-type glutamate receptors are recruited. acetaldehyde dehydrogenase (ALDH2; enzymes involved
Specically, the infralimbic medial prefrontal cortex-to- in the metabolism of alcohol) are reportedly protective
nucleus accumbens shell circuit recruits calcium- against alcoholism.160 Intriguingly, an evolutionarily
permeable AMPA receptors,144 and the prelimbic medial conserved GABA synthesis pathway mediated by aldehyde
prefrontal cortex-to-nucleus accumbens core circuit dehydrogenase 1a1 (ALDH1a1) has been identied in the
recruits non-calcium-permeable AMPA receptors. The ventral tegmental area.161 GABA co-release is modulated
blockade of metabotropic glutamate 1 receptors in the by binge-like doses of alcohol administered to mice,
nucleus accumbens blocks cue-induced incubation and diminished ALDH1a1 leads to enhanced alcohol
reinstatement and cocaine-primed reinstatement; as a consumption and preference. Similarly, polymorphisms
result, these receptors are potential therapeutic targets.145,146 in the genes for cytochrome P450 2A6 and 2D6 (enzymes
www.thelancet.com/psychiatry Vol 3 August 2016 767
Review
involved in nicotine and opioid metabolism, respectively) during adolescence reduced the density of myelinated
are reportedly protective against nicotine addiction.162 axons in the medial prefrontal cortex.176
From the perspective of the withdrawal/negative aect Environmental factors that have been consistently
stage, in humans, two single-nucleotide polymorphisms associated with the propensity to self-administer drugs
in the CRF1 receptor gene (CRHR1) were associated with include structural factors (eg, low socioeconomic status
binge drinking in adolescents and excessive drinking in and associated lack of social support systems), proximal
adults.163 Moreover, homozygosity at one of these single- factors (eg, parental drug use and dependence, as per the
nucleotide polymorphisms (rs1876831, C allele) was DSM-IV, poor quality of parenting, parental depression,
associated with heavy drinking in relation to stressful life and sibling and peer inuences), and more distal
events in adolescents.164 factors (eg, drug availability, school, neighbourhood
From the perspective of the preoccupation/anticipation characteristics, advertising, and the media).177,178 Stress is
stage, a genetic knock-in mouse that biologically also a common feature that increases the risk for drug
recapitulates a common human mutation in the gene for abuse. The mechanisms that are responsible for stress-
fatty acid amide hydrolase (C385A [Pro129Thr], rs324420) induced increases in vulnerability to drug use and relapse
presented a reduction of fatty acid amide hydrolase in those who are addicted are not yet well understood,
expression associated with the variant allele that selectively but evidence suggests that the stress-responsive
enhanced fronto-amygdala connectivity and fear extinction neuropeptide CRF is involved through its eects in the
learning and decreased anxiety-like behaviours; this result amygdala and hypothalamic-pituitary-adrenal axis.20
suggests another possible molecular genetic target for
neuroplasticity in the preoccupation/anticipation and Relevance to behavioural addictive disorders
withdrawal/negative aect stages.165 The three stages of the addiction cycle are pervasive and
Although such approaches do not guarantee that these form common domains in non-drug addictions, also
genes convey vulnerability in the human population, they known as process addictions,179 such as pathological
provide viable candidates for exploring the genetic basis of gambling, binge-eating disorder, compulsive buying, and
endophenotypes that are associated with addiction. internet addiction disorder.180 Non-drug addictions elaborate
self-regulation failures similarly to drug addictions, with
Developmental exposure as a key component of transitions from impulsivity to compulsivity and a chronic
vulnerability for drug and alcohol use disorders relapsing trajectory. Similar brain mechanisms, particularly
Normal developmental processes might result in higher with regard to reward decits, stress sensitisation, and
risk for drug use at some stages of the lifecycle than impaired inhibitory control, have been observed in
others. Experimentation, as well as the process of behavioural or process addictions, as well as decreases in
addiction, most often starts in adolescence,166 a period striatal dopamine D2 receptors associated with reduced
during which the brain undergoes important activity in prefrontal cortices,181 decits in the Go craving
developmental changes.167 Beginning in preadolescence system and Stop inhibition systems,182,183 and dysregulation
and continuing into the mid-20s, cortical grey matter of the stress axis.184
volumes reduce, which reects a normal pruning
process;168 white matter volume increases over the course Implications for medication development
of adolescence, reecting increases in connectivity, Our contention is that the eld of the neurobiology of
including axonal extension and myelination.168 Drug addiction has excellent and validated animal models and
exposure during adolescence is associated with more well developed clinical models that, combined with the
chronic and intensive use and greater risk of a substance neurocircuitry analysis herein, will provide a unique
use disorder than is initiation at older ages.167,169172 Normal approach to medication development that emphasises
adolescent-specic behaviours (eg, risk-taking, novelty- excessive incentive salience, the loss of brain reward
seeking, and high sensitivity to peer pressure) increase function, and the gain of stress function that drive
the propensity to experiment with legal and illegal negative reinforcement (ie, the dark side of addiction) and
drugs,167 which might reect the incomplete development the dysregulation of executive function, all of which are
of brain regions (eg, myelination of frontal lobe regions) key components that drive compulsive drug seeking.13,185
that are involved in the processes of executive control and Advances in diagnosis that reect such an endophenotype
motivation.168 Heavy alcohol use during adolescence is or research domain criterion approach will synergistically
associated with a range of neurobehavioural sequelae, combine with advances in neurobiology to promote novel
including impairments in visuospatial processing, pharmacotherapeutic, brain stimulation (ie, transcranial
attention, and memory,173,174 and adolescents who had magnetic stimulation and direct electrical stimulation),
engaged in episodes of heavy drinking presented faster and behavioural treatments.186
declining volumes in some neocortical grey matter
regions and smaller increases in regional white matter Conclusions and future directions
volumes than did non-drinking adolescents.175 We elaborate here a heuristic framework based on the
Additionally, voluntary binge drinking in animal models behavioural and imaging phenotypes of addiction as three
768 www.thelancet.com/psychiatry Vol 3 August 2016
Review
7 Moeller FG, Barratt ES, Dougherty DM, Schmitz JM, Swann AC.
Search strategy and selection criteria Psychiatric aspects of impulsivity. Am J Psychiatry 2001; 158: 178393.
8 Berlin GS, Hollander E. Compulsivity, impulsivity, and the DSM-5
We identied seminal articles published in peer-reviewed process. CNS Spectr 2014; 19: 6268.
journals and reports that were pertinent to the neurobiology 9 Koob GF, Volkow ND. Neurocircuitry of addiction.
of addiction using in-house expertise, consultations with Neuropsychopharmacology 2010; 35: 21738.
other experts in the eld, and searches of key databases, 10 Shippenberg TS, Koob GF. Recent advances in animal models of
drug addiction and alcoholism. In: Davis KL, Charney D, Coyle JT,
including PubMed. The exclusion and inclusion criteria for the Nemero C, eds. Neuropsychopharmacology: the fth generation of
articles were deliberately kept exible. The scope of the progress. Philadelphia: Lippincott Williams and Wilkins,
review was expanded based on ndings from the review of 2002: 138197.
11 Geyer MA, Markou A. Animal models of psychiatric disorders.
key papers and reports. In: Bloom FE, Kupfer DJ, eds. Psychopharmacology: the fourth
generation of progress. New York: Raven Press, 1995: 78798.
12 Koob GF, Le Moal M. Drug addiction, dysregulation of reward, and
allostasis. Neuropsychopharmacology 2001; 24: 97129.
stages linked by three functional domains that are 13 Koob GF, Mason BJ. Existing and future drugs for the treatment of
mediated by three major neurobiological circuits (basal the dark side of addiction. Annu Rev Pharmacol Toxicol 2016;
ganglia, extended amygdala, and prefrontal cortex) and 56: 299322.
14 Goldstein RZ, Volkow ND. Dysfunction of the prefrontal cortex in
numerous microcircuits of neuroplasticity. We outline addiction: neuroimaging ndings and clinical implications.
18 neurochemically dened mini circuits that can Nat Rev Neurosci 2011; 12: 65269.
independently or interactively load the outputs of the 15 Everitt BJ, Belin D, Economidou D, Pelloux Y, Dalley JW,
major common neurobiological circuits to produce Robbins TW. Review. Neural mechanisms underlying the
vulnerability to develop compulsive drug-seeking habits and
incentive salience and compulsive-like habits, negative addiction. Philos Trans R Soc Lond B Biol Sci 2008; 363: 312535.
emotional states of low reward and excessive stress, and 16 Mitchell JM, ONeil JP, Janabi M, Marks SM, Jagust WJ, Fields HL.
compromised executive function. Identication of these Alcohol consumption induces endogenous opioid release in the
human orbitofrontal cortex and nucleus accumbens. Sci Transl Med
molecular and neurochemical loads on the circuitry 2012; 4: 116ra6.
provides key information about vulnerability, resilience, 17 Cunningham KA, Anastasio NC. Serotonin at the nexus of
treatment, and recovery from addiction, as well as impulsivity and cue reactivity in cocaine addiction.
Neuropharmacology 2014; 76: 46078.
information on how dierent drugs of abuse enter the 18 Vashchinkina E, Panhelainen A, Aitta-Aho T, Korpi ER. GABAA
overall addiction cycle. Non-drug addictions, such as receptor drugs and neuronal plasticity in reward and aversion: focus
pathological gambling, have a similar behavioural on the ventral tegmental area. Front Pharmacol 2014; 5: 256.
phenotype that ts into the three-stage cycle. Preliminary 19 Dani JA, Heinemann S. Molecular and cellular aspects of nicotine
abuse. Neuron 1996; 16: 90508.
imaging data also suggest common neurobiology. Key 20 Koob GF, Buck CL, Cohen A, et al. Addiction as a stress surfeit
questions that remain are what genetic factors load these disorder. Neuropharmacology 2014; 76: 37082.
mini circuits, how the environment conveys epigenetic 21 Carlezon WA Jr, Nestler EJ, Neve RL. Herpes simplex virus-mediated
gene transfer as a tool for neuropsychiatric research.
inuences on these circuits, and how these circuits recover Crit Rev Neurobiol 2000; 14: 4767.
or do not recover with abstinence and treatment. Resolving 22 Delfs JM, Zhu Y, Druhan JP, Aston-Jones G. Noradrenaline in the
these questions should provide biomarkers for prevention, ventral forebrain is critical for opiate withdrawal-induced aversion.
Nature 2000; 403: 43034.
behavioural windows of development for prevention, novel
23 Boutrel B, de Lecea L. Addiction and arousal: the hypocretin
behavioural and pharmaceutical treatments, and ultimately connection. Physiol Behav 2008; 93: 94751.
a neurobiological understanding of recovery. 24 Maldonado R, Girdlestone D, Roques BP. RP 67580, a selective
antagonist of neurokinin-1 receptors, modies some of the
Contributors
naloxone-precipitated morphine withdrawal signs in rats.
Both authors contributed equally to writing the manuscript. Neurosci Lett 1993; 156: 13540.
Declaration of interests 25 Martinez D, Narendran R, Foltin RW, et al. Amphetamine-induced
We declare no competing interests. dopamine release: markedly blunted in cocaine dependence and
predictive of the choice to self-administer cocaine. Am J Psychiatry
Acknowledgments 2007; 164: 62229.
We thank Michael Arends for assistance with manuscript preparation. 26 Al-Hasani R, Bruchas MR. Molecular mechanisms of opioid
References receptor-dependent signaling and behavior. Anesthesiology 2011;
1 American Psychiatric Association. Diagnostic and statistical manual 115: 136381.
of mental disorders, 5th edn. Washington DC: American Psychiatric 27 Heilig M, Thorsell A. Brain neuropeptide Y (NPY) in stress and
Publishing, 2013. alcohol dependence. Rev Neurosci 2002; 13: 8594.
2 Koob GF, Le Moal M. Addiction and the brain antireward system. 28 Economidou D, Cippitelli A, Stopponi S, et al. Activation of brain
Annu Rev Psychol 2008; 59: 2953. NOP receptors attenuates acute and protracted alcohol withdrawal
3 McLellan AT, Lewis DC, OBrien CP, Kleber HD. Drug dependence, symptoms in the rat. Alcohol Clin Exp Res 2011; 35: 74755.
a chronic medical illness: implications for treatment, insurance, 29 Sidhpura N, Parsons LH. Endocannabinoid-mediated synaptic
and outcomes evaluation. JAMA 2000; 284: 168995. plasticity and addiction-related behavior. Neuropharmacology 2011;
4 Koob GF, Le Moal M. Drug abuse: hedonic homeostatic 61: 107087.
dysregulation. Science 1997; 278: 5258. 30 Sarnyai Z, Kovcs GL. Role of oxytocin in the neuroadaptation to
5 Goldstein RZ, Volkow ND. Drug addiction and its underlying drugs of abuse. Psychoneuroendocrinology 1994; 19: 85117.
neurobiological basis: neuroimaging evidence for the involvement 31 Fotros A, Casey KF, Larcher K, et al. Cocaine cue-induced dopamine
of the frontal cortex. Am J Psychiatry 2002; 159: 164252. release in amygdala and hippocampus: a high-resolution PET [F]
6 Baumeister RF, Heatherton TF, Tice DM. Losing control: how and fallypride study in cocaine dependent participants.
why people fail at self-regulation. San Diego: Academic Press, 1994. Neuropsychopharmacology 2013; 38: 178088.
www.thelancet.com/psychiatry Vol 3 August 2016 769
Review
32 Kalivas PW. The glutamate homeostasis hypothesis of addiction. 58 Ungless MA, Whistler JL, Malenka RC, Bonci A. Single cocaine
Nat Rev Neurosci 2009; 10: 56172. exposure in vivo induces long-term potentiation in dopamine
33 Shalev U, Grimm JW, Shaham Y. Neurobiology of relapse to heroin neurons. Nature 2001; 411: 58387.
and cocaine seeking: a review. Pharmacol Rev 2002; 54: 142. 59 Belin D, Everitt BJ. Cocaine seeking habits depend upon
34 Wise RA. Roles for nigrostriatalnot just mesocorticolimbic dopamine-dependent serial connectivity linking the ventral with the
dopamine in reward and addiction. Trends Neurosci 2009; 32: 51724. dorsal striatum. Neuron 2008; 57: 43241.
35 Volkow ND, Wang GJ, Telang F, et al. Profound decreases in 60 Roitman MF, Stuber GD, Phillips PE, Wightman RM, Carelli RM.
dopamine release in striatum in detoxied alcoholics: possible Dopamine operates as a subsecond modulator of food seeking.
orbitofrontal involvement. J Neurosci 2007; 27: 1270006. J Neurosci 2004; 24: 126571.
36 Volkow ND, Fowler JS, Wang GJ. The addicted human brain: 61 Volkow ND, Tomasi D, Wang GJ, et al. Stimulant-induced
insights from imaging studies. J Clin Invest 2003; 111: 144451. dopamine increases are markedly blunted in active cocaine abusers.
37 Caine SB, Thomsen M, Gabriel KI, et al. Lack of self-administration Mol Psychiatry 2014; 19: 103743.
of cocaine in dopamine D1 receptor knock-out mice. J Neurosci 2007; 62 Grace AA. The tonic/phasic model of dopamine system regulation
27: 1314050. and its implications for understanding alcohol and psychostimulant
38 Zweifel LS, Parker JG, Lobb CJ, et al. Disruption of NMDAR- craving. Addiction 2000; 95 (suppl 2): S11928.
dependent burst ring by dopamine neurons provides selective 63 Volkow ND, Morales M. The brain on drugs: from reward to
assessment of phasic dopamine-dependent behavior. addiction. Cell 2015; 162: 71225.
Proc Natl Acad Sci USA 2009; 106: 728188. 64 Bello EP, Mateo Y, Gelman DM, et al. Cocaine supersensitivity and
39 Norman AB, Tabet MR, Norman MK, Fey BK, Tsibulsky VL, enhanced motivation for reward in mice lacking dopamine D2
Millard RW. The anity of D2-like dopamine receptor antagonists autoreceptors. Nat Neurosci 2011; 14: 103338.
determines the time to maximal eect on cocaine self- 65 Vorel SR, Ashby CR Jr, Paul M, et al. Dopamine D3 receptor
administration. J Pharmacol Exp Ther 2011; 338: 72428. antagonism inhibits cocaine-seeking and cocaine-enhanced brain
40 Caine SB, Negus SS, Mello NK, et al. Role of dopamine D2-like reward in rats. J Neurosci 2002; 22: 9595603.
receptors in cocaine self-administration: studies with D2 receptor 66 Koob GF, Le Moal M. Plasticity of reward neurocircuitry and the
mutant mice and novel D2 receptor antagonists. J Neurosci 2002; dark side of drug addiction. Nat Neurosci 2005; 8: 144244.
22: 297788. 67 Garavan H, Pankiewicz J, Bloom A, et al. Cue-induced cocaine
41 Durieux PF, Bearzatto B, Guiducci S, et al. D2R striatopallidal craving: neuroanatomical specicity for drug users and drug
neurons inhibit both locomotor and drug reward processes. stimuli. Am J Psychiatry 2000; 157: 178998.
Nat Neurosci 2009; 12: 39395. 68 Koob GF, Bloom FE. Cellular and molecular mechanisms of drug
42 Covey DP, Roitman MF, Garris PA. Illicit dopamine transients: dependence. Science 1988; 242: 71523.
reconciling actions of abused drugs. Trends Neurosci 2014; 37: 20010. 69 Weiss F, Markou A, Lorang MT, Koob GF. Basal extracellular
43 Lscher C, Malenka RC. Drug-evoked synaptic plasticity in dopamine levels in the nucleus accumbens are decreased during
addiction: from molecular changes to circuit remodeling. Neuron cocaine withdrawal after unlimited-access self-administration.
2011; 69: 65063. Brain Res 1992; 593: 31418.
44 Tolu S, Eddine R, Marti F, et al. Co-activation of VTA DA and GABA 70 Volkow ND, Wang GJ, Fowler JS, et al. Decreased striatal
neurons mediates nicotine reinforcement. Mol Psychiatry 2013; dopaminergic responsiveness in detoxied cocaine-dependent
18: 38293. subjects. Nature 1997; 386: 83033.
45 Belin D, Jonkman S, Dickinson A, Robbins TW, Everitt BJ. 71 Stinus L, Le Moal M, Koob GF. Nucleus accumbens and amygdala
Parallel and interactive learning processes within the basal ganglia: are possible substrates for the aversive stimulus eects of opiate
relevance for the understanding of addiction. Behav Brain Res 2009; withdrawal. Neuroscience 1990; 37: 76773.
199: 89102. 72 Minkowski CP, Epstein D, Frost JJ, Gorelick DA. Dierential
46 Wolf ME, Ferrario CR. AMPA receptor plasticity in the nucleus response to IV carfentanil in chronic cocaine users and healthy
accumbens after repeated exposure to cocaine. controls. Addict Biol 2012; 17: 14955.
Neurosci Biobehav Rev 2010; 35: 185211. 73 Davidson M, Shanley B, Wilce P. Increased NMDA-induced
47 Grieder TE, Herman MA, Contet C, et al. VTA CRF neurons excitability during ethanol withdrawal: a behavioural and
mediate the aversive eects of nicotine withdrawal and promote histological study. Brain Res 1995; 674: 9196.
intake escalation. Nat Neurosci 2014; 17: 175158. 74 Dahchour A, De Witte P, Bolo N, et al. Central eects of
48 Fowler CD, Lu Q, Johnson PM, Marks MJ, Kenny PJ. Habenular 5 acamprosate: part 1. Acamprosate blocks the glutamate increase in
nicotinic receptor subunit signalling controls nicotine intake. the nucleus accumbens microdialysate in ethanol withdrawn rats.
Nature 2011; 471: 597601. Psychiatry Res 1998; 82: 10714.
49 Heilig M. The NPY system in stress, anxiety and depression. 75 Piazza PV, Le Moal ML. Pathophysiological basis of vulnerability to
Neuropeptides 2004; 38: 21324. drug abuse: role of an interaction between stress, glucocorticoids,
50 George O, Sanders C, Freiling J, et al. Recruitment of medial and dopaminergic neurons. Annu Rev Pharmacol Toxicol 1996;
prefrontal cortex neurons during alcohol withdrawal predicts 36: 35978.
cognitive impairment and excessive alcohol drinking. 76 Walker BM, Koob GF. Pharmacological evidence for a motivational
Proc Natl Acad Sci USA 2012; 109: 1815661. role of -opioid systems in ethanol dependence.
51 Vorel SR, Liu X, Hayes RJ, Spector JA, Gardner EL. Relapse to Neuropsychopharmacology 2008; 33: 64352.
cocaine-seeking after hippocampal theta burst stimulation. Science 77 Schlosburg JE, Whiteld TW Jr, Park PE, et al. Long-term
2001; 292: 117578. antagonism of opioid receptors prevents escalation of and
52 Everitt BJ, Wolf ME. Psychomotor stimulant addiction: a neural increased motivation for heroin intake. J Neurosci 2013; 33: 1938492.
systems perspective. J Neurosci 2002; 22: 331220. 78 Whiteld TW Jr, Schlosburg JE, Wee S, et al. opioid receptors in
53 Shaham Y, Shalev U, Lu L, De Wit H, Stewart J. The reinstatement the nucleus accumbens shell mediate escalation of
model of drug relapse: history, methodology and major ndings. methamphetamine intake. J Neurosci 2015; 35: 4296305.
Psychopharmacology (Berl) 2003; 168: 320. 79 Hikosaka O. The habenula: from stress evasion to value-based
54 Naqvi NH, Bechara A. The hidden island of addiction: the insula. decision-making. Nat Rev Neurosci 2010; 11: 50313.
Trends Neurosci 2009; 32: 5667. 80 Matsumoto M, Hikosaka O. Lateral habenula as a source of
55 Robbins TW. Relationship between reward-enhancing and negative reward signals in dopamine neurons. Nature 2007;
stereotypical eects of psychomotor stimulant drugs. Nature 1976; 447: 111115.
264: 5759. 81 Kimura M, Satoh T, Matsumoto N. What does the habenula tell
56 Berridge MJ. Calcium signalling remodelling and disease. dopamine neurons? Nat Neurosci 2007; 10: 67778.
Biochem Soc Trans 2012; 40: 297309. 82 Salas R, Sturm R, Boulter J, De Biasi M. Nicotinic receptors in the
57 Schultz W, Dayan P, Montague PR. A neural substrate of prediction habenulo-interpeduncular system are necessary for nicotine
and reward. Science 1997; 275: 159399. withdrawal in mice. J Neurosci 2009; 29: 301418.
770 www.thelancet.com/psychiatry Vol 3 August 2016
Review
83 Gillan CM, Morein-Zamir S, Urcelay GP, et al. Enhanced avoidance 105 Jasinska AJ, Stein EA, Kaiser J, Naumer MJ, Yalachkov Y.
habits in obsessive-compulsive disorder. Biol Psychiatry 2014; Factors modulating neural reactivity to drug cues in addiction:
75: 63138. a survey of human neuroimaging studies. Neurosci Biobehav Rev
84 Reisiger AR, Kauing J, Manzoni O, Cador M, Georges F, Caill S. 2014; 38: 116.
Nicotine self-administration induces CB1-dependent LTP in the bed 106 Volkow ND, Wang GJ, Telang F, et al. Cocaine cues and dopamine
nucleus of the stria terminalis. J Neurosci 2014; 34: 428592. in dorsal striatum: mechanism of craving in cocaine addiction.
85 Gunduz-Cinar O, MacPherson KP, Cinar R, et al. Convergent J Neurosci 2006; 26: 658388.
translational evidence of a role for anandamide in amygdala- 107 Milella MS, Fotros A, Gravel P, et al. Cocaine cue-induced
mediated fear extinction, threat processing and stress-reactivity. dopamine release in the human prefrontal cortex.
Mol Psychiatry 2013; 18: 81323. J Psychiatry Neurosci 2016; 41: 150207.
86 Hirvonen J, Goodwin RS, Li CT, et al. Reversible and regionally 108 Volkow ND, Wang GJ, Fowler JS, et al. Association of
selective downregulation of brain cannabinoid CB1 receptors in methylphenidate-induced craving with changes in right striato-
chronic daily cannabis smokers. Mol Psychiatry 2012; 17: 64249. orbitofrontal metabolism in cocaine abusers: implications in
87 Ceccarini J, Hompes T, Verhaeghen A, et al. Changes in cerebral addiction. Am J Psychiatry 1999; 156: 1926.
CB1 receptor availability after acute and chronic alcohol abuse and 109 Volkow ND, Wang GJ, Fowler JS, Tomasi D, Telang F. Addiction:
monitored abstinence. J Neurosci 2014; 34: 282231. beyond dopamine reward circuitry. Proc Natl Acad Sci USA 2011;
88 Leventhal AM, Kahler CW, Ray LA, et al. Anhedonia and 108: 1503742.
amotivation in psychiatric outpatients with fully remitted stimulant 110 Rando K, Hong KI, Bhagwagar Z, et al. Association of frontal and
use disorder. Am J Addict 2008; 17: 21823. posterior cortical gray matter volume with time to alcohol relapse:
89 Tiany ST, Carter BL, Singleton EG. Challenges in the a prospective study. Am J Psychiatry 2011; 168: 18392.
manipulation, assessment and interpretation of craving relevant 111 Bonson KR, Grant SJ, Contoreggi CS, et al. Neural systems and
variables. Addiction 2000; 95 (suppl 2): S17787. cue-induced cocaine craving. Neuropsychopharmacology 2002;
90 Geisler S, Wise RA. Functional implications of glutamatergic 26: 37686.
projections to the ventral tegmental area. Rev Neurosci 2008; 112 Pelchat ML, Johnson A, Chan R, Valdez J, Ragland JD. Images of
19: 22744. desire: food-craving activation during fMRI. Neuroimage 2004;
91 Koob GF, Arends MA, Le Moal M. Drugs, addiction, and the brain. 23: 148693.
San Diego: Academic Press, 2014. 113 Wang Z, Faith M, Patterson F, et al. Neural substrates of abstinence-
92 Jentsch JD, Taylor JR. Impulsivity resulting from frontostriatal induced cigarette cravings in chronic smokers. J Neurosci 2007;
dysfunction in drug abuse: implications for the control of behavior 27: 1403540.
by reward-related stimuli. Psychopharmacology (Berl) 1999; 114 Naqvi NH, Rudrauf D, Damasio H, Bechara A. Damage to the
146: 37390. insula disrupts addiction to cigarette smoking. Science 2007;
93 Chudasama Y, Robbins TW. Functions of frontostriatal systems in 315: 53134.
cognition: comparative neuropsychopharmacological studies in 115 Clark L, Bechara A, Damasio H, Aitken MR, Sahakian BJ,
rats, monkeys and humans. Biol Psychol 2006; 73: 1938. Robbins TW. Dierential eects of insular and ventromedial
94 McFarland K, Kalivas PW. The circuitry mediating cocaine-induced prefrontal cortex lesions on risky decision-making. Brain 2008;
reinstatement of drug-seeking behavior. J Neurosci 2001; 131: 131122.
21: 865563. 116 Verdejo-Garcia A, Contreras-Rodrguez O, Fonseca F, et al.
95 Vanderschuren LJMJ, Di Ciano P, Everitt BJ. Involvement of the Functional alteration in frontolimbic systems relevant to moral
dorsal striatum in cue-controlled cocaine seeking. J Neurosci 2005; judgment in cocaine-dependent subjects. Addict Biol 2014;
25: 866570. 19: 27281.
96 Shaham Y, Higheld D, Delfs J, Leung S, Stewart J. Clonidine 117 Snchez MM, Young LJ, Plotsky PM, Insel TR. Autoradiographic
blocks stress-induced reinstatement of heroin seeking in rats: and in situ hybridization localization of corticotropin-releasing
an eect independent of locus coeruleus noradrenergic neurons. factor 1 and 2 receptors in nonhuman primate brain. J Comp Neurol
Eur J Neurosci 2000; 12: 292302. 1999; 408: 36577.
97 Shaham Y, Erb S, Leung S, Buczek Y, Stewart J. CP-154,526, a 118 Goudriaan AE, de Ruiter MB, van den Brink W, Oosterlaan J,
selective, non-peptide antagonist of the corticotropin-releasing Veltman DJ. Brain activation patterns associated with cue
factor1 receptor attenuates stress-induced relapse to drug seeking in reactivity and craving in abstinent problem gamblers, heavy
cocaine- and heroin-trained rats. Psychopharmacology (Berl) 1998; smokers and healthy controls: an fMRI study. Addict Biol 2010;
137: 18490. 15: 491503.
98 Merlo Pich E, Lorang M, Yeganeh M, et al. Increase of extracellular 119 Janes AC, Pizzagalli DA, Richardt S, et al. Brain reactivity to
corticotropin-releasing factor-like immunoreactivity levels in the smoking cues prior to smoking cessation predicts ability to
amygdala of awake rats during restraint stress and ethanol withdrawal maintain tobacco abstinence. Biol Psychiatry 2010; 67: 72229.
as measured by microdialysis. J Neurosci 1995; 15: 543947. 120 Volkow ND, Fowler JS. Addiction, a disease of compulsion and
99 De Witte P, Littleton J, Parot P, Koob G. Neuroprotective and drive: involvement of the orbitofrontal cortex. Cereb Cortex 2000;
abstinence-promoting eects of acamprosate: elucidating the 10: 31825.
mechanism of action. CNS Drugs 2005; 19: 51737. 121 Volkow ND, Wang GJ, Fowler JS, Telang F. Overlapping neuronal
100 Valdez GR, Roberts AJ, Chan K, et al. Increased ethanol circuits in addiction and obesity: evidence of systems pathology.
self-administration and anxiety-like behavior during acute ethanol Philos Trans R Soc Lond B Biol Sci 2008a; 363: 3191200.
withdrawal and protracted abstinence: regulation by 122 Volkow ND, Fowler JS, Wang GJ, Baler R, Telang F. Imaging
corticotropin-releasing factor. Alcohol Clin Exp Res 2002; 26: 1494501. dopamines role in drug abuse and addiction. Neuropharmacology
101 Kober H, Lacadie CM, Wexler BE, Malison RT, Sinha R, 2009; 56 (suppl 1): 38.
Potenza MN. Brain activity during cocaine craving and gambling 123 Volkow ND, Fowler JS, Wang GJ, et al. Decreased dopamine D2
urges: an fMRI study. Neuropsychopharmacology 2016; 41: 62837. receptor availability is associated with reduced frontal metabolism
102 Lee JH, Lim Y, Wiederhold BK, Graham SJ. A functional magnetic in cocaine abusers. Synapse 1993; 14: 16977.
resonance imaging (FMRI) study of cue-induced smoking craving 124 Volkow ND, Chang L, Wang GJ, et al. Low level of brain dopamine
in virtual environments. Appl Psychophysiol Biofeedback 2005; D2 receptors in methamphetamine abusers: association with
30: 195204. metabolism in the orbitofrontal cortex. Am J Psychiatry 2001;
103 Risinger RC, Salmeron BJ, Ross TJ, et al. Neural correlates of high 158: 201521.
and craving during cocaine self-administration using BOLD fMRI. 125 Lee B, London ED, Poldrack RA, et al. Striatal dopamine d2/d3
Neuroimage 2005; 26: 1097108. receptor availability is reduced in methamphetamine dependence
104 Volkow ND, Wang GJ, Ma Y, et al. Activation of orbital and medial and is linked to impulsivity. J Neurosci 2009; 29: 1473440.
prefrontal cortex by methylphenidate in cocaine-addicted subjects 126 Wang GJ, Smith L, Volkow ND, et al. Decreased dopamine activity
but not in controls: relevance to addiction. J Neurosci 2005; predicts relapse in methamphetamine abusers. Mol Psychiatry 2012;
25: 393239. 17: 91825.
www.thelancet.com/psychiatry Vol 3 August 2016 771
Review
127 Niendam TA, Laird AR, Ray KL, Dean YM, Glahn DC, Carter CS. 151 Barak S, Liu F, Ben Hamida S, et al. Disruption of alcohol-related
Meta-analytic evidence for a superordinate cognitive control memories by mTORC1 inhibition prevents relapse. Nat Neurosci
network subserving diverse executive functions. 2013; 16: 111117.
Cogn Aect Behav Neurosci 2012; 12: 24168. 152 Lerman C, Berrettini W. Elucidating the role of genetic factors in
128 Contreras-Rodrguez O, Albein-Urios N, Perales JC, et al. Cocaine- smoking behavior and nicotine dependence.
specic neuroplasticity in the ventral striatum network is linked to Am J Med Genet B Neuropsychiatr Genet 2003; 118B: 4854.
delay discounting and drug relapse. Addiction 2015; 110: 195362. 153 Matthes HWD, Maldonado R, Simonin F, et al. Loss of
129 Bryant RA, Felmingham KL, Kemp AH, et al. Neural networks of morphine-induced analgesia, reward eect and withdrawal
information processing in posttraumatic stress disorder: a symptoms in mice lacking the -opioid-receptor gene. Nature
functional magnetic resonance imaging study. Biol Psychiatry 2005; 1996; 383: 81923.
58: 11118. 154 Ledent C, Valverde O, Cossu G, et al. Unresponsiveness to
130 Damasio AR. The somatic marker hypothesis and the possible cannabinoids and reduced addictive eects of opiates in CB1
functions of the prefrontal cortex. Philos Trans R Soc Lond B Biol Sci receptor knockout mice. Science 1999; 283: 40104.
1996; 351: 141320. 155 Laakso A, Mohn AR, Gainetdinov RR, Caron MG. Experimental
131 Bechara A, Damasio H, Damasio AR, Lee GP. Dierent genetic approaches to addiction. Neuron 2002; 36: 21328.
contributions of the human amygdala and ventromedial prefrontal 156 Uhl GR, Grow RW. The burden of complex genetics in brain
cortex to decision-making. J Neurosci 1999; 19: 547381. disorders. Arch Gen Psychiatry 2004; 61: 22329.
132 Johnstone T, van Reekum CM, Urry HL, Kalin NH, Davidson RJ. 157 Hancock DB, Reginsson GW, Gaddis NC, et al. Genome-wide
Failure to regulate: counterproductive recruitment of top-down meta-analysis reveals common splice site acceptor variant in
prefrontal-subcortical circuitry in major depression. J Neurosci 2007; CHRNA4 associated with nicotine dependence. Transl Psychiatr
27: 887784. 2015; 5: e651.
133 Pitman RK, Rasmusson AM, Koenen KC, et al. Biological studies of 158 Olfson E, Saccone NL, Johnson EO, et al. Rare, low frequency and
post-traumatic stress disorder. Nat Rev Neurosci 2012; 13: 76987. common coding variants in CHRNA5 and their contribution to
134 Mcdonald AJ, Mascagni F, Guo L. Projections of the medial and nicotine dependence in European and African Americans.
lateral prefrontal cortices to the amygdala: a Phaseolus vulgaris Mol Psychiatry 2016; 21: 60107.
leucoagglutinin study in the rat. Neuroscience 1996; 71: 5575. 159 Nelson EC, Agrawal A, Heath AC, et al. Evidence of CNIH3
135 Volkow ND, Fowler JS, Wang GJ, et al. Cognitive control of drug involvement in opioid dependence. Mol Psychiatry 2016; 21: 60814.
craving inhibits brain reward regions in cocaine abusers. 160 Chen YC, Lu RB, Peng GS, et al. Alcohol metabolism and
Neuroimage 2010; 49: 253643. cardiovascular response in an alcoholic patient homozygous for the
136 Self DW, Genova LM, Hope BT, Barnhart WJ, Spencer JJ, Nestler EJ. ALDH2*2 variant gene allele. Alcohol Clin Exp Res 1999; 23: 185360.
Involvement of cAMP-dependent protein kinase in the nucleus 161 Kim JI, Ganesan S, Luo SX, et al. Aldehyde dehydrogenase 1a1
accumbens in cocaine self-administration and relapse of mediates a GABA synthesis pathway in midbrain dopaminergic
cocaine-seeking behavior. J Neurosci 1998; 18: 184859. neurons. Science 2015; 350: 10206.
137 Edwards S, Koob GF. Neurobiology of dysregulated motivational 162 Benowitz NL, Swan GE, Jacob P 3rd, Lessov-Schlaggar CN,
systems in drug addiction. Future Neurol 2010; 5: 393401. Tyndale RF. CYP2A6 genotype and the metabolism and disposition
138 Carlezon WA Jr, Thome J, Olson VG, et al. Regulation of cocaine kinetics of nicotine. Clin Pharmacol Ther 2006; 80: 45767.
reward by CREB. Science 1998; 282: 227275. 163 Treutlein J, Kissling C, Frank J, et al. Genetic association of the
139 Nestler EJ. Molecular mechanisms of drug addiction. human corticotropin releasing hormone receptor 1 (CRHR1) with
Neuropharmacology 2004; 47 (suppl 1): 2432. binge drinking and alcohol intake patterns in two independent
140 Pandey SC, Sakharkar AJ, Tang L, Zhang H. Potential role of samples. Mol Psychiatry 2006; 11: 594602.
adolescent alcohol exposure-induced amygdaloid histone 164 Blomeyer D, Treutlein J, Esser G, Schmidt MH, Schumann G,
modications in anxiety and alcohol intake during adulthood. Laucht M. Interaction between CRHR1 gene and stressful life
Neurobiol Dis 2015; 82: 60719. events predicts adolescent heavy alcohol use. Biol Psychiatry 2008;
141 Nestler EJ. Is there a common molecular pathway for addiction? 63: 14651.
Nat Neurosci 2005; 8: 144549. 165 Dincheva I, Drysdale AT, Hartley CA, et al. FAAH genetic variation
142 Kelz MB, Chen J, Carlezon WA Jr, et al. Expression of the enhances fronto-amygdala function in mouse and human.
transcription factor deltaFosB in the brain controls sensitivity to Nat Commun 2015; 6: 6395.
cocaine. Nature 1999; 401: 27276. 166 Wagner FA, Anthony JC. From rst drug use to drug dependence;
143 Feng J, Wilkinson M, Liu X, et al. Chronic cocaine-regulated developmental periods of risk for dependence upon marijuana,
epigenomic changes in mouse nucleus accumbens. Genome Biol cocaine, and alcohol. Neuropsychopharmacology 2002; 26: 47988.
2014; 15: R65. 167 Spear LP. Adolescent neurodevelopment. J Adolesc Health 2013;
144 Conrad KL, Tseng KY, Uejima JL, et al. Formation of accumbens 52 (suppl 2): S713.
GluR2-lacking AMPA receptors mediates incubation of cocaine 168 Giedd JN, Blumenthal J, Jeries NO, et al. Brain development
craving. Nature 2008; 454: 11821. during childhood and adolescence: a longitudinal MRI study.
145 Schmidt HD, Kimmey BA, Arreola AC, Pierce RC. Group I Nat Neurosci 1999; 2: 86163.
metabotropic glutamate receptor-mediated activation of PKC in 169 Grant BF, Harford TC, Muthen BO. Age at onset of alcohol use and
the nucleus accumbens core promotes the reinstatement of cocaine DSM-IV alcohol abuse and dependence: a 12-year follow-up study.
seeking. Addict Biol 2015; 20: 28596. J Subst Abuse 2001; 13: 493304.
146 Loweth JA, Scheyer AF, Milovanovic M, et al. Synaptic depression 170 Volkow ND. Altered pathways: drug abuse and age of onset.
via mGluR1 positive allosteric modulation suppresses cue-induced Addict Professional 2006; 26: 29.
cocaine craving. Nat Neurosci 2014; 17: 7380. 171 Hingson RW, Heeren T, Winter MR. Age of alcohol-dependence
147 Costa-Mattioli M, Sossin WS, Klann E, Sonenberg N. Translational onset: associations with severity of dependence and seeking
control of long-lasting synaptic plasticity and memory. Neuron 2009; treatment. Pediatrics 2006; 118: e75563.
61: 1026. 172 King KM, Chassin L. A prospective study of the eects of age of
148 Costa-Mattioli M, Monteggia LM. mTOR complexes in initiation of alcohol and drug use on young adult substance
neurodevelopmental and neuropsychiatric disorders. Nat Neurosci dependence. J Stud Alcohol Drugs 2007; 68: 25665.
2013; 16: 153743. 173 Hanson KL, Medina KL, Padula CB, Tapert SF, Brown SA.
149 Neasta J, Barak S, Hamida SB, Ron D. mTOR complex 1: a key Impact of adolescent alcohol and drug use on neuropsychological
player in neuroadaptations induced by drugs of abuse. J Neurochem functioning in young adulthood: 10-year outcomes.
2014; 130: 17284. J Child Adolesc Subst Abuse 2011; 20: 13554.
150 Shi X, Miller JS, Harper LJ, Poole RL, Gould TJ, Unterwald EM. 174 Squeglia LM, Spadoni AD, Infante MA, Myers MG, Tapert SF.
Reactivation of cocaine reward memory engages the Akt/GSK3/ Initiating moderate to heavy alcohol use predicts changes in
mTOR signaling pathway and can be disrupted by GSK3 inhibition. neuropsychological functioning for adolescent girls and boys.
Psychopharmacology (Berl) 2014; 231: 310918. Psychol Addict Behav 2009; 23: 71522.
772 www.thelancet.com/psychiatry Vol 3 August 2016
Review
175 Squeglia LM, Tapert SF, Sullivan EV, et al. Brain development in 182 Yau YH, Potenza MN. Gambling disorder and other behavioral
heavy-drinking adolescents. Am J Psychiatry 2015; 172: 53142. addictions: recognition and treatment. Harv Rev Psychiatry 2015;
176 Vargas WM, Bengston L, Gilpin NW, Whitcomb BW, 23: 13446.
Richardson HN. Alcohol binge drinking during adolescence or 183 Volkow ND, Baler RD. NOW vs LATER brain circuits: implications
dependence during adulthood reduces prefrontal myelin in male for obesity and addiction. Trends Neurosci 2015; 38: 34552.
rats. J Neurosci 2014; 34: 1477782. 184 Lemieux A, alAbsi M. Stress psychobiology in the context of
177 Hawkins JD, Catalano RF, Miller JY. Risk and protective factors for addiction medicine: from drugs of abuse to behavioral addictions.
alcohol and other drug problems in adolescence and early Prog Brain Res 2016; 223: 4362.
adulthood: implications for substance abuse prevention. 185 Koob GF, Kenneth Lloyd G, Mason BJ. Development of
Psychol Bull 1992; 112: 64105. pharmacotherapies for drug addiction: a Rosetta stone approach.
178 Sloboda Z, Glantz MD, Tarter RE. Revisiting the concepts of risk Nat Rev Drug Discov 2009; 8: 50015.
and protective factors for understanding the etiology and 186 Kwako LE, Momenan R, Litten RZ, Koob GF, Goldman D.
development of substance use and substance use disorders: Addictions neuroclinical assessment: a neuroscience-based
implications for prevention. Subst Use Misuse 2012; 47: 94462. framework for addictive disorders. Biol Psychiatry 2015; published
179 Smith DE. The process of addictions and the new ASAM denition online Nov 17. DOI:10.1016/j.biopsych.2015.10.024
of addiction. J Psychoactive Drugs 2012; 44: 14.
180 Koob GF. Impulse control disorders. San Diego: Cognella, 2013.
181 Volkow ND, Wang GJ, Telang F, et al. Low dopamine striatal D2
receptors are associated with prefrontal metabolism in obese
subjects: possible contributing factors. Neuroimage 2008;
42: 153743.
www.thelancet.com/psychiatry Vol 3 August 2016 773
You might also like
- Addiction Neuroethics: The Ethics of Addiction Neuroscience Research and TreatmentFrom EverandAddiction Neuroethics: The Ethics of Addiction Neuroscience Research and TreatmentNo ratings yet
- Neurobiología AdicciónDocument14 pagesNeurobiología AdicciónClaudia Araneda JorqueraNo ratings yet
- 02 Neurobiologia de La AdiccionDocument25 pages02 Neurobiologia de La AdiccionZaira Camila CriolloNo ratings yet
- Circuitos de La AdiccionDocument28 pagesCircuitos de La AdiccionJose Andres AlvarezNo ratings yet
- Biological Substrates of Addiction 2014Document30 pagesBiological Substrates of Addiction 2014Jacky KarNo ratings yet
- The Neurobiology of Addictive DisordersDocument8 pagesThe Neurobiology of Addictive DisordersDee DeeNo ratings yet
- The New England Journal of Medicine Vol 374Document16 pagesThe New England Journal of Medicine Vol 374tatih meilaniNo ratings yet
- Neurobiology of Addiction: Toward The Development of New TherapiesDocument16 pagesNeurobiology of Addiction: Toward The Development of New Therapiesguessimfloatin GNo ratings yet
- Chapter 2 NeurobiologyDocument31 pagesChapter 2 Neurobiologyanitha1118No ratings yet
- The Darkside of Drug AddictionDocument3 pagesThe Darkside of Drug Addictioninetwork2001100% (1)
- Drug Addiction: The Neurobiology of Disrupted Self-Control: Ruben D. Baler and Nora D. VolkowDocument8 pagesDrug Addiction: The Neurobiology of Disrupted Self-Control: Ruben D. Baler and Nora D. VolkowDavid James PNo ratings yet
- Volkow (2015) - Biomarkers in Substance UseDocument4 pagesVolkow (2015) - Biomarkers in Substance UseJulieht RodriguezNo ratings yet
- Addiction - 2005 - Spanagel - Addiction and Its Brain ScienceDocument10 pagesAddiction - 2005 - Spanagel - Addiction and Its Brain ScienceGliter CorpuzNo ratings yet
- Smith 2014Document8 pagesSmith 2014Dilan GutierrezNo ratings yet
- Addiction: Steven N TreistmanDocument7 pagesAddiction: Steven N TreistmanFrancisco BecerraNo ratings yet
- DopamineDocument11 pagesDopaminecaioaccorsiNo ratings yet
- The Neurobiology of AddictionDocument24 pagesThe Neurobiology of AddictionErnesto Jaramillo ValenzuelaNo ratings yet
- 1 Neuroscience of Drug Reward and AddictionDocument26 pages1 Neuroscience of Drug Reward and Addictionasdrubal5martinezNo ratings yet
- The Brain Disease Model of Addiction: Butler Center For Research May 2021Document2 pagesThe Brain Disease Model of Addiction: Butler Center For Research May 2021cultura84No ratings yet
- Understanding PDFDocument4 pagesUnderstanding PDFchyntiaNo ratings yet
- Mecanismos Neurobiológicos de La Adicción A DrogasDocument18 pagesMecanismos Neurobiológicos de La Adicción A DrogasJuan Antonio Herrera LealNo ratings yet
- Adiccion PDFDocument22 pagesAdiccion PDFAnonymous Csk4UVPONo ratings yet
- Alfa AgonistasDocument7 pagesAlfa AgonistasSumroachNo ratings yet
- Theories of Addiction: By-Manali Naphade (MA Psychology Sem III)Document24 pagesTheories of Addiction: By-Manali Naphade (MA Psychology Sem III)Manali NaphadeNo ratings yet
- The Neurobiological Underpinnings of AddictionDocument9 pagesThe Neurobiological Underpinnings of AddictionShaun Shelly100% (1)
- Mclaughlin Revised Literature ReviewDocument15 pagesMclaughlin Revised Literature Reviewapi-625527319No ratings yet
- Alcohol, Opioids, and Other SubstanceDocument98 pagesAlcohol, Opioids, and Other SubstanceKesha PatelNo ratings yet
- Neurobehavior & Special Sense System Tutor's Guide Substance Related DisorderDocument22 pagesNeurobehavior & Special Sense System Tutor's Guide Substance Related DisorderNovi AdriNo ratings yet
- Garrison (2014) - Neuroimaging and Biomarkers in Addiction Treatment.Document9 pagesGarrison (2014) - Neuroimaging and Biomarkers in Addiction Treatment.Julieht RodriguezNo ratings yet
- The Role of Cognitive Control in Cocaine DependenceDocument9 pagesThe Role of Cognitive Control in Cocaine Dependenceanon_708494042No ratings yet
- Senior Project - Is Drug Adicction A Brain Disease-2Document13 pagesSenior Project - Is Drug Adicction A Brain Disease-2api-668828236No ratings yet
- The Neurocircuitry of Addiction: An Overview: ReviewDocument14 pagesThe Neurocircuitry of Addiction: An Overview: ReviewHarrison JessimerNo ratings yet
- Ajp 2007 164 1 4Document3 pagesAjp 2007 164 1 4Maritsa EngelbrechtNo ratings yet
- Addiction DisordersDocument32 pagesAddiction Disordersgideon gideonNo ratings yet
- Lewis, 2017Document12 pagesLewis, 2017Angela JimenezNo ratings yet
- Neurobiología de Las AdiccionesDocument17 pagesNeurobiología de Las AdiccionesYasmin IquiseNo ratings yet
- The Overlapping Neurobiology of Addiction and ADHDDocument3 pagesThe Overlapping Neurobiology of Addiction and ADHDDanilo Borges Dos SantosNo ratings yet
- Neurobiological Basis of Drug Reward and Reinforcement: Chapter OutlineDocument18 pagesNeurobiological Basis of Drug Reward and Reinforcement: Chapter Outlineal sharpNo ratings yet
- Smoke Cessation 1Document15 pagesSmoke Cessation 1Ricardo GalhardoniNo ratings yet
- Drug Addiction PDFDocument16 pagesDrug Addiction PDFHeinz FischerNo ratings yet
- Addiction and The Brain Development Not DiseaseDocument13 pagesAddiction and The Brain Development Not DiseaseGayneOrtegaNo ratings yet
- Addiction: A Dysregulation of Satiety and Inflammatory ProcessesDocument27 pagesAddiction: A Dysregulation of Satiety and Inflammatory ProcessesAulas EspañolNo ratings yet
- SPP 03 1 13Document10 pagesSPP 03 1 13Zenith xNo ratings yet
- Pharmacological Treatments For Cocaine Craving - What Is The Way Forward - A Systematic ReviewDocument46 pagesPharmacological Treatments For Cocaine Craving - What Is The Way Forward - A Systematic ReviewBianca CmrnNo ratings yet
- Volkow 2018Document12 pagesVolkow 2018Marcos AlexandreNo ratings yet
- Uso de Sustancias y EsquizofreniaDocument10 pagesUso de Sustancias y Esquizofreniaracm89No ratings yet
- Can Biophysical Models Give Insight Into The Synaptic Changes Associated With Addiction?Document19 pagesCan Biophysical Models Give Insight Into The Synaptic Changes Associated With Addiction?dhananjay.dagaNo ratings yet
- Use Drugs - Pay To Die Drugs AddictionDocument11 pagesUse Drugs - Pay To Die Drugs AddictionelvineNo ratings yet
- Neuroscience of Substance Abuse and DependenceDocument62 pagesNeuroscience of Substance Abuse and Dependencedrkadiyala2100% (1)
- Drugs Behavior 8th Hancock Test BankDocument15 pagesDrugs Behavior 8th Hancock Test BankSaifoqqNo ratings yet
- Psychedelic: TherapyDocument7 pagesPsychedelic: TherapyzoeNo ratings yet
- Understanding The Disease of AddictionDocument7 pagesUnderstanding The Disease of AddictionJoão MaiaNo ratings yet
- A Communication Process of EMBA Faculty of Business Studites at Jahangirnar UniversityDocument2 pagesA Communication Process of EMBA Faculty of Business Studites at Jahangirnar UniversitySourav Nath Shaan 1611875630No ratings yet
- 0-Addicto Craving Hone-Blanchet (2014)Document15 pages0-Addicto Craving Hone-Blanchet (2014)gildas5d5almeidaNo ratings yet
- The Neurobiology of Impulsivity and Substance Use Disorders Implications For TreatmentDocument33 pagesThe Neurobiology of Impulsivity and Substance Use Disorders Implications For TreatmentLanaNo ratings yet
- Understanding Drug Abuse and AddictionDocument4 pagesUnderstanding Drug Abuse and AddictionAdam ChabchoubNo ratings yet
- Acetylsistein Journal 1Document13 pagesAcetylsistein Journal 1Arifa LainaNo ratings yet
- Addiction Recovery Through Photovoice Qualitative StudyDocument18 pagesAddiction Recovery Through Photovoice Qualitative Studylapkas donjuanNo ratings yet
- 1 s2.0 S0079612315001909 MainDocument27 pages1 s2.0 S0079612315001909 Mainqnx22605No ratings yet
- Psychopharmacological Treatment of Obsessive-Compulsive Disorder (OCD)Document27 pagesPsychopharmacological Treatment of Obsessive-Compulsive Disorder (OCD)godolfintorentNo ratings yet
- Cannabis MigraineDocument6 pagesCannabis MigraineMarinaNo ratings yet
- Castano Et AlDocument20 pagesCastano Et AlMarinaNo ratings yet
- Tobacco Prevention Education in Schools For The Deaf: The Faculty PerspectiveDocument25 pagesTobacco Prevention Education in Schools For The Deaf: The Faculty PerspectiveMarinaNo ratings yet
- Effects of Adolescent Marijuana UseDocument10 pagesEffects of Adolescent Marijuana UseMarinaNo ratings yet
- 6 Clinical - Health - Psychology - and - Primary - Care - Practical - Advice - and - Clinical - Guidance - For - Successful - Collaboration PDFDocument260 pages6 Clinical - Health - Psychology - and - Primary - Care - Practical - Advice - and - Clinical - Guidance - For - Successful - Collaboration PDFMarinaNo ratings yet
- Follow-Up Study Longterm UnemploymentDocument9 pagesFollow-Up Study Longterm UnemploymentMarinaNo ratings yet
- Trauma-Related Impairment in Children-A Survey in Sri Lankan Provinces Affected by Armed ConflictDocument9 pagesTrauma-Related Impairment in Children-A Survey in Sri Lankan Provinces Affected by Armed ConflictMarinaNo ratings yet
- The Long-Term Effects of War Experiences On Children's Depression in The Republic of CroatiaDocument13 pagesThe Long-Term Effects of War Experiences On Children's Depression in The Republic of CroatiaMarinaNo ratings yet
- Benefits Associated With FamilyDocument7 pagesBenefits Associated With FamilyMarinaNo ratings yet
- Family Dinner Meal FrequencyDocument9 pagesFamily Dinner Meal FrequencyMarinaNo ratings yet
- Vegetarian EatingDocument8 pagesVegetarian EatingMarinaNo ratings yet
- Compliance With Mediterranean DietDocument6 pagesCompliance With Mediterranean DietMarinaNo ratings yet
- Preliminary Report On A New Aversion Therapy For Male HomosexualsDocument3 pagesPreliminary Report On A New Aversion Therapy For Male HomosexualsMarinaNo ratings yet
- BJP 2008 Lock 426Document3 pagesBJP 2008 Lock 426MarinaNo ratings yet
- Health Concern Questionnaire PT CurrentDocument14 pagesHealth Concern Questionnaire PT CurrentMarinaNo ratings yet
- African-American Adolescent DietDocument226 pagesAfrican-American Adolescent DietMarinaNo ratings yet
- Aversion Therapy Issues: A Note of Clarification: University of Oregon, Eugene, Oregon 97403Document5 pagesAversion Therapy Issues: A Note of Clarification: University of Oregon, Eugene, Oregon 97403MarinaNo ratings yet
- 1 s2.0 074054729090033M MainDocument9 pages1 s2.0 074054729090033M MainMarinaNo ratings yet
- English Icd10Document3 pagesEnglish Icd10MarinaNo ratings yet
- Aversion by Fiat: The Problem of "Face Validity" in Behavior TherapyDocument7 pagesAversion by Fiat: The Problem of "Face Validity" in Behavior TherapyMarinaNo ratings yet
- Case Historles AND Shorter CoimmunlcationsDocument4 pagesCase Historles AND Shorter CoimmunlcationsMarinaNo ratings yet
- Financial Crises Impact On Mental Health and Suggested Responses.Document6 pagesFinancial Crises Impact On Mental Health and Suggested Responses.MarinaNo ratings yet
- General HealthDocument2 pagesGeneral HealthMarinaNo ratings yet
- Evidence For The 2008 Economic Crisis Exacerbating Depression in Hong KongDocument9 pagesEvidence For The 2008 Economic Crisis Exacerbating Depression in Hong KongMarinaNo ratings yet
- Economic Crises and Mortality - A Review of The LiteratureDocument8 pagesEconomic Crises and Mortality - A Review of The LiteratureMarinaNo ratings yet
- Clinicians' Reports of The Impact of The 2008 Financial Crisis On Mental Health Clients.Document23 pagesClinicians' Reports of The Impact of The 2008 Financial Crisis On Mental Health Clients.MarinaNo ratings yet
- Economic Crisis, Restrictive Policies, and The Population's Health and Health Care - The Greek Case.Document8 pagesEconomic Crisis, Restrictive Policies, and The Population's Health and Health Care - The Greek Case.MarinaNo ratings yet
- Hypothalamus: Pleasure&Reward, AversionDocument18 pagesHypothalamus: Pleasure&Reward, AversionJayalakshmi KNo ratings yet
- Neuroscience of Addiction Review: George F. Koob, Pietro Paolo Sanna, and Floyd E. BloomDocument10 pagesNeuroscience of Addiction Review: George F. Koob, Pietro Paolo Sanna, and Floyd E. Bloomadri90No ratings yet
- Hypothalamus: Pleasure&Reward, Aversion: Jayalakshmi.k M.Sc. Clinical PsychologyDocument19 pagesHypothalamus: Pleasure&Reward, Aversion: Jayalakshmi.k M.Sc. Clinical PsychologyJayalakshmi KNo ratings yet
- Neurochemistry of MusicDocument15 pagesNeurochemistry of MusicLisa CerezoNo ratings yet
- EmotionDocument56 pagesEmotionvalcobacl.681.studNo ratings yet
- DopamineDocument28 pagesDopamineNTA UGC-NETNo ratings yet
- NPB 168 Practice Exam 2Document5 pagesNPB 168 Practice Exam 2Kim Wong100% (1)
- Neurobiology of SchizophreniaDocument54 pagesNeurobiology of SchizophreniaPriyash JainNo ratings yet
- Littrell - What Neurobiology Has To SayDocument19 pagesLittrell - What Neurobiology Has To SayRosalyn S. LeeNo ratings yet
- Scientific American, A Division of Nature America, Inc. Scientific AmericanDocument13 pagesScientific American, A Division of Nature America, Inc. Scientific AmericanXiomara Velázquez LandaNo ratings yet
- CH 15 Drug Use Addiction Brains Reward CircuitsDocument11 pagesCH 15 Drug Use Addiction Brains Reward CircuitsNatalia KarasiewiczNo ratings yet
- Neurotransmitters and Psychiatry PDFDocument22 pagesNeurotransmitters and Psychiatry PDFcapriciousbelal93% (14)
- Brain Circuit Dysfunction in Post - Traumatic Stress Disorder: From Mouse To Man - Robert J. FensterDocument17 pagesBrain Circuit Dysfunction in Post - Traumatic Stress Disorder: From Mouse To Man - Robert J. FensterAlvaro Arboledas LoriteNo ratings yet
- 10 - Donahoe. Burgos, & Palmer (1993) PDFDocument24 pages10 - Donahoe. Burgos, & Palmer (1993) PDFMiguel100% (1)
- Alcoholism PINKAN PDFDocument68 pagesAlcoholism PINKAN PDFMartga Bella RahimiNo ratings yet
- Male and Female Sexual Dysfunction-072343266XDocument287 pagesMale and Female Sexual Dysfunction-072343266XLikoh Timothy100% (2)
- Your Brain in Love. by Fisher H.Document5 pagesYour Brain in Love. by Fisher H.doraNo ratings yet
- 2023 - Atlas of Sleep Medicine, 3rd Edition - (Robert Joseph Thomas, Sushanth Bhat Etc.)Document797 pages2023 - Atlas of Sleep Medicine, 3rd Edition - (Robert Joseph Thomas, Sushanth Bhat Etc.)César EscobarNo ratings yet
- P3000 Complete and Peace Culture 2019Document111 pagesP3000 Complete and Peace Culture 2019maria iliana simionNo ratings yet
- Neurobiology of Emotions: An Update: International Review of PsychiatryDocument16 pagesNeurobiology of Emotions: An Update: International Review of PsychiatryEva Bastida MercadoNo ratings yet
- The Biologic Basis For Libido: James G. Pfaus, PHD, and Lisa A.Scepkowski, MaDocument6 pagesThe Biologic Basis For Libido: James G. Pfaus, PHD, and Lisa A.Scepkowski, MaPaul AsturbiarisNo ratings yet
- Limbic SystemDocument49 pagesLimbic SystemBaishali DebNo ratings yet
- Behavioural AddictionsDocument15 pagesBehavioural AddictionsnishthasainiNo ratings yet
- NeuropsychologyDocument76 pagesNeuropsychologybushra231100% (1)
- Cannabis Use and Cannabis Use DisorderDocument24 pagesCannabis Use and Cannabis Use DisorderRafael RibeiroNo ratings yet
- A Clinical Guide To Pediatric SleepDocument304 pagesA Clinical Guide To Pediatric Sleepchucrallah1968No ratings yet
- Disorders of Diminished Motivation: Robert S. Marin, MD Patricia A. Wilkosz, MD, PHDDocument13 pagesDisorders of Diminished Motivation: Robert S. Marin, MD Patricia A. Wilkosz, MD, PHDAnanda WulandariNo ratings yet
- BDSM ArticlesDocument31 pagesBDSM ArticlesPrincezed19887171No ratings yet
- Opioid Pharmacology and Pharmaco Genetics DraftDocument18 pagesOpioid Pharmacology and Pharmaco Genetics DraftPrudhvi Sai NidadavoluNo ratings yet
- Neurobiology of Addiction PDFDocument6 pagesNeurobiology of Addiction PDFAriol ZereNo ratings yet