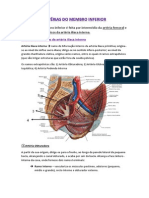
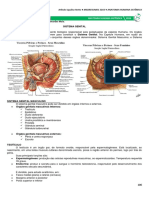
You might also like
- Medresumos 2014 - Anatomia Humana Sistêmica 03 - Sistema ArticularDocument8 pagesMedresumos 2014 - Anatomia Humana Sistêmica 03 - Sistema ArticularGabriel HansNo ratings yet
- Medresumos 2014 - Anatomia Humana Sistêmica 06 - Sistema CirculatórioDocument11 pagesMedresumos 2014 - Anatomia Humana Sistêmica 06 - Sistema CirculatórioGabriel HansNo ratings yet
- 07 - Tecido Cartilaginoso PDFDocument4 pages07 - Tecido Cartilaginoso PDFShâmara Stéfany Souza GuimarãesNo ratings yet
- Medresumos 2014 - Anatomia Humana Sistêmica 01 - IntroduçãoDocument9 pagesMedresumos 2014 - Anatomia Humana Sistêmica 01 - IntroduçãoGabriel HansNo ratings yet
- MiologiaDocument141 pagesMiologiaArthur Sacramento100% (10)
- Irrigação membro inferior artérias ilíaca femoral poplíteaDocument13 pagesIrrigação membro inferior artérias ilíaca femoral poplíteaTamára Marçal100% (1)
- 03 - Núcleo InterfásicoDocument2 pages03 - Núcleo InterfásicoForwomenNo ratings yet
- Cílios e flagelos: estrutura e funçãoDocument7 pagesCílios e flagelos: estrutura e funçãoForwomenNo ratings yet
- 05 - Tecido Adiposo PDFDocument2 pages05 - Tecido Adiposo PDFShâmara Stéfany Souza Guimarães100% (2)
- Tecido ÓsseoDocument6 pagesTecido ÓsseoaeadaNo ratings yet
- Ciclo celular, fases da mitose e sistema de controleDocument5 pagesCiclo celular, fases da mitose e sistema de controleForwomenNo ratings yet
- Glandulas Endocrinas e ExocrinasDocument10 pagesGlandulas Endocrinas e Exocrinaspolyannamaramendonca100% (3)
- Sistema genital masculinoDocument14 pagesSistema genital masculinoForwomenNo ratings yet
- Peptídeos e proteínas: insulina, glucagon e outrosDocument6 pagesPeptídeos e proteínas: insulina, glucagon e outrosJéssica FrancoNo ratings yet
- Complexo de Golgi: estrutura e funçõesDocument5 pagesComplexo de Golgi: estrutura e funçõesForwomenNo ratings yet
- Introdução aos tecidos animaisDocument17 pagesIntrodução aos tecidos animaisCaroline PetrollNo ratings yet
- 15 - Aparelho Circulatório - MedresumoDocument6 pages15 - Aparelho Circulatório - MedresumoaeadaNo ratings yet
- Medresumos 2014 - Anatomia Humana Sistêmica 04 - Sistema MuscularDocument17 pagesMedresumos 2014 - Anatomia Humana Sistêmica 04 - Sistema MuscularGabriel HansNo ratings yet
- β-oxidação de lipídiosDocument44 pagesβ-oxidação de lipídiosMaria Eduarda Frota100% (1)
- Tecido Epitelial ResumoDocument6 pagesTecido Epitelial ResumoAlex BertoNo ratings yet
- Neuroanatomia Faculdade Lisboa 2007/2008Document343 pagesNeuroanatomia Faculdade Lisboa 2007/2008Joao Jose R100% (1)
- Tecido ósseo: características, funções e histologiaDocument38 pagesTecido ósseo: características, funções e histologiaAudrey RosolenNo ratings yet
- Sistema DigestorioDocument28 pagesSistema DigestorioDany JunqueiraNo ratings yet
- Aula 8-Tecido MuscularDocument45 pagesAula 8-Tecido Musculartelesdan1944No ratings yet
- Sistema muscular: estrutura e contraçãoDocument18 pagesSistema muscular: estrutura e contraçãoGessica Fabiana Cardoso0% (1)
- LeucinoseDocument2 pagesLeucinoseBárbara CostaNo ratings yet
- Estrutura e funções das paredes abdominaisDocument23 pagesEstrutura e funções das paredes abdominaisAna Carolina NonatoNo ratings yet
- Fisiologia III 01 - Neurofisiologia - Completa - Med Resumos (Fev-2012)Document0 pagesFisiologia III 01 - Neurofisiologia - Completa - Med Resumos (Fev-2012)zuendson6737No ratings yet
- Célula Estrutura e Função MedresumosDocument251 pagesCélula Estrutura e Função MedresumosAndersonFelix50% (2)
- Embriologia Do Sistema CardiovascularDocument2 pagesEmbriologia Do Sistema CardiovascularElise Assunção100% (1)
- Bases da Oncologia: Diferenças entre Célula Normal e CâncerosaDocument9 pagesBases da Oncologia: Diferenças entre Célula Normal e CâncerosaLavínia LafetáNo ratings yet
- Músculo esquelético: estrutura e contraçãoDocument5 pagesMúsculo esquelético: estrutura e contraçãomariany100% (2)
- Histologia Tecido OsseoDocument5 pagesHistologia Tecido OsseoKlarissa LimaNo ratings yet
- Sistema EsqueléticoDocument8 pagesSistema EsqueléticoEva CarvalhoNo ratings yet
- 03 - Teníase e Cisticercose HumanaDocument7 pages03 - Teníase e Cisticercose HumanaDiego FonsecaNo ratings yet
- Resumo Bioquímica Ciclo Da UreiaDocument4 pagesResumo Bioquímica Ciclo Da UreiaRaíssa XimenesNo ratings yet
- NEUROANATOMIA 10 - Macroscopia Do Diencéfalo - MED RESUMOS 2012 PDFDocument3 pagesNEUROANATOMIA 10 - Macroscopia Do Diencéfalo - MED RESUMOS 2012 PDFdellionkingNo ratings yet
- Histologia HumanaDocument90 pagesHistologia HumanaJhefferson GleizerNo ratings yet
- Bio Qui MicaDocument21 pagesBio Qui MicaCarolayne Souza Pinto100% (1)
- Os ácidos nucleicos: DNA e RNADocument4 pagesOs ácidos nucleicos: DNA e RNAAYLA APARECIDA LORENA SEIXASNo ratings yet
- Fisiologia Humana SimuladoDocument8 pagesFisiologia Humana SimuladoMarcia Nogueira FuturanutricionistaNo ratings yet
- Esquema de Embriologia DigestórioDocument3 pagesEsquema de Embriologia Digestóriojorgecf2375% (4)
- MEDRESUMOS 2017 - CAC - Anatomia Topográfica IDocument69 pagesMEDRESUMOS 2017 - CAC - Anatomia Topográfica IGabriel EliasNo ratings yet
- Relatório de HistologiaDocument6 pagesRelatório de HistologiaSara BeatrizNo ratings yet
- Bioquimica MedresumoDocument102 pagesBioquimica MedresumoSilvio Raul BenitezNo ratings yet
- Síntese de lipídeos e esteróidesDocument3 pagesSíntese de lipídeos e esteróidesMary FreitasNo ratings yet
- Processamento de RNA mensageiro em eucariotasDocument3 pagesProcessamento de RNA mensageiro em eucariotasairton barrosNo ratings yet
- Sistema DigestórioDocument2 pagesSistema DigestórioBreno Gonçalves100% (1)
- 01 - Introdução À ParasitologiaDocument2 pages01 - Introdução À ParasitologiaRafaella AraujoNo ratings yet
- Aula 01 DNA, RNA, História Da Genética. Genomas.Document58 pagesAula 01 DNA, RNA, História Da Genética. Genomas.Davi ÁlefeNo ratings yet
- Embriologia Respiratório e CardiovascularDocument10 pagesEmbriologia Respiratório e CardiovascularVanessaNo ratings yet
- Questionario Av2 2021Document5 pagesQuestionario Av2 2021Letycia Fernandes100% (1)
- Embriologia do aparelho digestivoDocument23 pagesEmbriologia do aparelho digestivoAna Carolina100% (1)
- Desenvolvimento do sistema locomotorDocument71 pagesDesenvolvimento do sistema locomotorAndersonFelix100% (1)
- Resumo de Anatomia - Parede / Cavidade Abdominal e Aparelho DigestorioDocument47 pagesResumo de Anatomia - Parede / Cavidade Abdominal e Aparelho DigestorioLuiz Felipe100% (17)
- Tecido NervosoDocument12 pagesTecido NervosoVINICIUS SCARANELLO ROSSI100% (1)
- Histologia II - Sistema CirculatórioDocument131 pagesHistologia II - Sistema CirculatórioMaria MatiasNo ratings yet
- No caminho certo após uma cirurgia cardíaca: Um guia completo com orientações e recomendações para o pós-operatórioFrom EverandNo caminho certo após uma cirurgia cardíaca: Um guia completo com orientações e recomendações para o pós-operatórioNo ratings yet
- Voto Celso Mello Prisao LulaDocument6 pagesVoto Celso Mello Prisao LulaForwomenNo ratings yet
- Diagnóstico e tratamento de aneurismas da aorta abdominalDocument28 pagesDiagnóstico e tratamento de aneurismas da aorta abdominalForwomenNo ratings yet
- Diretrizes Atencao Saude Ocular InfanciaDocument42 pagesDiretrizes Atencao Saude Ocular InfanciaFelipe Dias100% (1)
- Diagnóstico e tratamento de aneurismas da aorta abdominalDocument28 pagesDiagnóstico e tratamento de aneurismas da aorta abdominalForwomenNo ratings yet
- Febrasgo - 11 - Assistencia Ao Trabalho de PartoDocument12 pagesFebrasgo - 11 - Assistencia Ao Trabalho de PartoEduardo SantosNo ratings yet
- Diretrizes sobre abordagem geral da hipertensão arterialDocument24 pagesDiretrizes sobre abordagem geral da hipertensão arterialGrace PelicioniNo ratings yet
- Doenca de Alzheimer-DiagnosticoDocument24 pagesDoenca de Alzheimer-DiagnosticoAnny Kelly BorgesNo ratings yet
- 0066 782X Abc 106 04 s2 0001Document35 pages0066 782X Abc 106 04 s2 0001ForwomenNo ratings yet
- Atualização Da Diretriz Brasileira de Dislipidemias e Prevenção Da Aterosclerose - 2017Document92 pagesAtualização Da Diretriz Brasileira de Dislipidemias e Prevenção Da Aterosclerose - 2017Saulo YuriNo ratings yet
- Abuso e dependência de álcoolDocument36 pagesAbuso e dependência de álcoolDiego Oliveira BritoNo ratings yet
- Tuberculose - Biosseguranca E Risco OcupacionalDocument17 pagesTuberculose - Biosseguranca E Risco OcupacionalFidel Alcolea BittencourtNo ratings yet
- Diretrizes Clínicas AsmaDocument18 pagesDiretrizes Clínicas AsmaDio JoseNo ratings yet
- 15370Document2 pages15370ForwomenNo ratings yet
- Uso Dos Anti-Inflamatorios Nao Hormonais Na Dor Cronica em Pacientes Com Osteoartrite (Osteoartrose) PDFDocument25 pagesUso Dos Anti-Inflamatorios Nao Hormonais Na Dor Cronica em Pacientes Com Osteoartrite (Osteoartrose) PDFForwomenNo ratings yet
- Diretrizes sobre abordagem geral da hipertensão arterialDocument24 pagesDiretrizes sobre abordagem geral da hipertensão arterialGrace PelicioniNo ratings yet
- CI 02 - Min SaúdeDocument3 pagesCI 02 - Min SaúdeForwomenNo ratings yet
- 351 Avaliaç - o Do Bem-Estar FetalDocument5 pages351 Avaliaç - o Do Bem-Estar FetalForwomenNo ratings yet
- Guia de Vigilância em SaúdeDocument706 pagesGuia de Vigilância em SaúdeRosalina Braga100% (1)
- Saude Crianca Nutricao To AlimentacaoDocument112 pagesSaude Crianca Nutricao To AlimentacaoJu KinjoNo ratings yet
- Arboviroses AhmDocument8 pagesArboviroses AhmForwomenNo ratings yet
- Manejo Clínicojan2018Document155 pagesManejo Clínicojan2018ForwomenNo ratings yet
- Casos Clínico 2 DengueDocument3 pagesCasos Clínico 2 DengueForwomenNo ratings yet
- Arbo Sit Epid Plano Jan 2018Document54 pagesArbo Sit Epid Plano Jan 2018ForwomenNo ratings yet
- Plano de Contingência Ubs Organização de ServiçosDocument27 pagesPlano de Contingência Ubs Organização de ServiçosForwomenNo ratings yet
- Fluxograma para Atendimento de Caso Suspeito de Dengue RevisadoDocument1 pageFluxograma para Atendimento de Caso Suspeito de Dengue RevisadoForwomenNo ratings yet
- Manejo Clínicojan2018Document155 pagesManejo Clínicojan2018ForwomenNo ratings yet
- Plano de Contingência UbsDocument7 pagesPlano de Contingência UbsForwomenNo ratings yet
- Plano de Contingência Ubs Organização de ServiçosDocument27 pagesPlano de Contingência Ubs Organização de ServiçosForwomenNo ratings yet
- Casos Clínico 1 Dengue1Document2 pagesCasos Clínico 1 Dengue1ForwomenNo ratings yet
- Caso clínico de paciente com dengue que evoluiu para óbitoDocument3 pagesCaso clínico de paciente com dengue que evoluiu para óbitoForwomenNo ratings yet
- Compilado Fisiologia e Biofisica (Unidade 01, 02, 03 e 04 Produzido Por Henrique RochaDocument185 pagesCompilado Fisiologia e Biofisica (Unidade 01, 02, 03 e 04 Produzido Por Henrique RochaRenata SousaNo ratings yet
- Morte celular: necrose e apoptoseDocument49 pagesMorte celular: necrose e apoptoseprietoml50% (2)
- 104 Hemograma Canino - Acima de 8 AnosDocument1 page104 Hemograma Canino - Acima de 8 AnospaubonfillgassosfrinfertortosaNo ratings yet
- Biologia - Fotossíntese e Respiração CelularDocument54 pagesBiologia - Fotossíntese e Respiração CelularFabio Tomas de AquinoNo ratings yet
- Manual Biologia UFCD1Document25 pagesManual Biologia UFCD1Ana Mafalda Torres100% (2)
- Microbiologia e Imunologia BacterianaDocument44 pagesMicrobiologia e Imunologia BacterianaTarcizio FilhoNo ratings yet
- Exercícios de Revisão BiocelDocument115 pagesExercícios de Revisão BiocelMaria Rosa DmengeonNo ratings yet
- Lista Atividades Lipideos e MembranasDocument9 pagesLista Atividades Lipideos e MembranasNATALIE OLIVEIRA DUARTENo ratings yet
- Anticorpos e Antigenos PerguntasDocument7 pagesAnticorpos e Antigenos Perguntasmaria eduardaNo ratings yet
- BioeletrogeneseDocument62 pagesBioeletrogeneseBeatriz CupolilloNo ratings yet
- Bioquímica-Fisiológica-Exame 210129 010644Document7 pagesBioquímica-Fisiológica-Exame 210129 010644Joana Carolina Teixeira RibeiroNo ratings yet
- Proteínas 2 2Document57 pagesProteínas 2 2Isabela RochaNo ratings yet
- Reprodução CelularDocument4 pagesReprodução Celular485trtrNo ratings yet
- Trabalho 2 de Biologia 1º AnoDocument2 pagesTrabalho 2 de Biologia 1º AnoCharles RibeiroNo ratings yet
- Exercícios Sobre Células Procariontes e EucariontesDocument3 pagesExercícios Sobre Células Procariontes e EucariontesRuiMarcoBicalhoNo ratings yet
- Aula 13Document36 pagesAula 13Nuno G. PintoNo ratings yet
- Regulacao Da Expressao GenicaDocument33 pagesRegulacao Da Expressao GenicaFelipe Alves de OliveiraNo ratings yet
- FecundaçãoDocument8 pagesFecundaçãoMaria SantosNo ratings yet
- Aula 4 - Parte A MutaçãoDocument35 pagesAula 4 - Parte A MutaçãoSamara RodriguesNo ratings yet
- CitologiaDocument45 pagesCitologiaNikilitosNo ratings yet
- Aula 10 Imunologia Dos TranslantesDocument35 pagesAula 10 Imunologia Dos TranslantesNENZIM FFNo ratings yet
- Obtenção de Matéria Pelos Seres Heterotróficos - ResumoDocument3 pagesObtenção de Matéria Pelos Seres Heterotróficos - Resumofrancisca soaresNo ratings yet
- Capitulo 1 Vitelogenese Nos Ecdizoa - Nematoides e Insetos Como Exemplos PDFDocument45 pagesCapitulo 1 Vitelogenese Nos Ecdizoa - Nematoides e Insetos Como Exemplos PDFJose Carlos Pelielo MattosNo ratings yet
- Bioquimica Aplicada A Enfermagem PDFDocument5 pagesBioquimica Aplicada A Enfermagem PDFpatrfrf100% (1)
- Biologia CelularDocument83 pagesBiologia CelularAmanda CarolineNo ratings yet
- 6 Imunidade Inata #SANARDocument26 pages6 Imunidade Inata #SANARodontouefs2020.1No ratings yet
- Prova Parcial A - GabaritoDocument3 pagesProva Parcial A - GabaritoSheila Souza VieiraNo ratings yet
- Imunologia - Aula 4Document14 pagesImunologia - Aula 4Valeria CasiqueNo ratings yet
- Mitose e MeioseDocument6 pagesMitose e Meioseabreuu50No ratings yet
- Introdução à Fisiologia CelularDocument83 pagesIntrodução à Fisiologia CelularDaniela 1616No ratings yet