You might also like
- Lecture03 NotesDocument4 pagesLecture03 NotesRoshan ShresthaNo ratings yet
- Project 2: Formatting and Processing Data Into Usable FormatsDocument1 pageProject 2: Formatting and Processing Data Into Usable FormatsRoshan ShresthaNo ratings yet
- Project 1: More Linux and The ShellDocument4 pagesProject 1: More Linux and The ShellRoshan ShresthaNo ratings yet
- Density of StatesDocument1 pageDensity of StatesRoshan ShresthaNo ratings yet
- Weinberg 2003Finally, learn something about the history of science,or at a minimum the history of your own branch of science. The least important reason for this is that the history may actually be of some use to you in your own scientific work. For instance, now and then scientists are hampered by believing one of the oversimplified models of science that have been proposed by philosophers from Francis Bacon to Thomas Kuhn and Karl Popper. The best antidote to the philosophy ofDocument1 pageWeinberg 2003Finally, learn something about the history of science,or at a minimum the history of your own branch of science. The least important reason for this is that the history may actually be of some use to you in your own scientific work. For instance, now and then scientists are hampered by believing one of the oversimplified models of science that have been proposed by philosophers from Francis Bacon to Thomas Kuhn and Karl Popper. The best antidote to the philosophy ofJonatan Vignatti MuñozNo ratings yet
- Sinc 2Document1 pageSinc 2Roshan ShresthaNo ratings yet
- Lecture08 NotesDocument4 pagesLecture08 NotesRoshan ShresthaNo ratings yet
- Introduction To UnixDocument31 pagesIntroduction To UnixRoshan ShresthaNo ratings yet
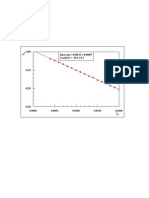
- 0.01000 Intercept 0.00197 0.00023 Gradient - 4.58 0.02Document1 page0.01000 Intercept 0.00197 0.00023 Gradient - 4.58 0.02Roshan ShresthaNo ratings yet
- 0.00 Intercept 0.00515 0.00067 Gradient - 10.6 0.1Document1 page0.00 Intercept 0.00515 0.00067 Gradient - 10.6 0.1Roshan ShresthaNo ratings yet
- 0.00 Intercept 0.00366 0.00027 Gradient - 8.06 0.02Document1 page0.00 Intercept 0.00366 0.00027 Gradient - 8.06 0.02Roshan ShresthaNo ratings yet
- 0.00 Intercept 0.00585 0.00048 Gradient - 11.4 0.0Document1 page0.00 Intercept 0.00585 0.00048 Gradient - 11.4 0.0Roshan ShresthaNo ratings yet
- Sec1 StatDescriptionDocument14 pagesSec1 StatDescriptionRoshan ShresthaNo ratings yet
- 0.00 Intercept 0.00326 0.00031 Gradient - 6.47 0.02Document1 page0.00 Intercept 0.00326 0.00031 Gradient - 6.47 0.02Roshan ShresthaNo ratings yet
- 0.00 Intercept 0.00382 0.00048 Gradient - 9.41 0.04Document1 page0.00 Intercept 0.00382 0.00048 Gradient - 9.41 0.04Roshan ShresthaNo ratings yet
- Sec2 MaxwellDocument8 pagesSec2 MaxwellRoshan ShresthaNo ratings yet
- Practical 1-To Constuct CE Amplifier For The Determination of Its Volatge GainDocument1 pagePractical 1-To Constuct CE Amplifier For The Determination of Its Volatge GainRoshan ShresthaNo ratings yet
- 2011 Exam Paper PDFDocument8 pages2011 Exam Paper PDFRoshan ShresthaNo ratings yet
- Homework 1Document5 pagesHomework 1tylermw334No ratings yet
- Free Lect Ron ModelDocument6 pagesFree Lect Ron ModelŞahin AktaşNo ratings yet
- Classical MechanicsDocument6 pagesClassical MechanicsRoshan ShresthaNo ratings yet
- GRE Flash Card Corrections Oct 2014Document1 pageGRE Flash Card Corrections Oct 2014Roshan ShresthaNo ratings yet
- Complex Analysis Part6Document7 pagesComplex Analysis Part6Roshan ShresthaNo ratings yet
- Difference AmplifierDocument21 pagesDifference AmplifierRoshan ShresthaNo ratings yet
- Preparation For The First Term Exam of First SemesterDocument1 pagePreparation For The First Term Exam of First SemesterRoshan ShresthaNo ratings yet
- 26 11 2013 014Document1 page26 11 2013 014Roshan ShresthaNo ratings yet
- 001124Document41 pages001124Ashish DhimanNo ratings yet
- Natural Background Sources: Cosmic RadiationDocument1 pageNatural Background Sources: Cosmic RadiationRoshan ShresthaNo ratings yet
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5784)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (890)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (72)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)