You might also like
- GRASTRITISDocument21 pagesGRASTRITISCamila Valentina HuamaniNo ratings yet
- Ulcera GastricaDocument10 pagesUlcera GastricaBetsaida Xiadani Díaz MondragónNo ratings yet
- Marco Teórico GastritisDocument10 pagesMarco Teórico GastritisGABRIELA YUCRA GUZMANNo ratings yet
- Ulcera PépticaDocument8 pagesUlcera PépticaKarla TNNo ratings yet
- Parasitologia QDDocument82 pagesParasitologia QDDario Gutiérrez QuirozNo ratings yet
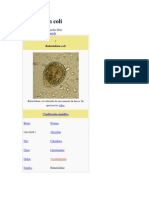
- Balantidiosis: infección por Balantidium coliDocument3 pagesBalantidiosis: infección por Balantidium coliDaniel GutiérrezNo ratings yet
- ParacitologiaDocument13 pagesParacitologiaDario Gutiérrez QuirozNo ratings yet
- El Infiltrado Inflamatorio Crónico Está Compuesto Fundamentalmente Por Linfocitos y PlasmocitosDocument4 pagesEl Infiltrado Inflamatorio Crónico Está Compuesto Fundamentalmente Por Linfocitos y Plasmocitoscrt200030% (1)
- Balantidiosis: Balantidium coli causa diarrea y estreñimientoDocument10 pagesBalantidiosis: Balantidium coli causa diarrea y estreñimientoMaria Celeste Abella MercadoNo ratings yet
- Medio de Keister Modificado para El Cultivo de Giardia Lamblia in Vitro ULTIMODocument8 pagesMedio de Keister Modificado para El Cultivo de Giardia Lamblia in Vitro ULTIMOSarahí D TrujilloNo ratings yet
- Helicobacter PyloriDocument5 pagesHelicobacter PyloriFredy Miguel CTNo ratings yet
- Helicobacter PyloriDocument11 pagesHelicobacter PyloriSara GNo ratings yet
- Resumen de Examen de ParasitologiaDocument49 pagesResumen de Examen de ParasitologiaAngel Luis FelizNo ratings yet
- De Helicobacter Pylori OfiDocument10 pagesDe Helicobacter Pylori OfiFreddy Ademar Plata MamaniNo ratings yet
- Balantidium ColiDocument8 pagesBalantidium ColiMaho PerezNo ratings yet
- Parasitosis IntestinalesDocument24 pagesParasitosis IntestinalesFrancisco ChalenNo ratings yet
- Atlas Benja 2 2Document27 pagesAtlas Benja 2 2andresdominguezbeltran1bNo ratings yet
- CiliadosDocument14 pagesCiliadosMARIA VELASQUEZ TORRESNo ratings yet
- H.pylori: generalidadesDocument7 pagesH.pylori: generalidadesjavierNo ratings yet
- Ppts Helicobacter PyloriDocument25 pagesPpts Helicobacter PyloriAngelNo ratings yet
- Balantidiosis o BalantidiasisDocument4 pagesBalantidiosis o BalantidiasisWuilliam ApaezNo ratings yet
- GastroenterologiaDocument15 pagesGastroenterologiaDay NuñezNo ratings yet
- Eap H.pyloriDocument6 pagesEap H.pyloriJonathan Juarez GarciaNo ratings yet
- Expo GastritisDocument57 pagesExpo GastritisDiana Cervantes100% (2)
- MARCO TEORICO-paDocument11 pagesMARCO TEORICO-paMaria TorresNo ratings yet
- AtlasDocument20 pagesAtlasEstuardo LavarredaNo ratings yet
- Helico B ActerDocument51 pagesHelico B ActerMirielis LoreNo ratings yet
- Importancia de Los Estudios Clínicos en El Diagnóstico de Enfermedades Patológicas Relacionadas Con Infecciones Intestinales. (H. Pylori)Document18 pagesImportancia de Los Estudios Clínicos en El Diagnóstico de Enfermedades Patológicas Relacionadas Con Infecciones Intestinales. (H. Pylori)priscila sanchez robleroNo ratings yet
- Patología benigna y neoplásica del estómagoDocument66 pagesPatología benigna y neoplásica del estómagoMaría AlejandrinaNo ratings yet
- Resumen MicroDocument7 pagesResumen MicroNeftaly CubiasNo ratings yet
- Balantidium coli: morfología, patogenia y diagnósticoDocument85 pagesBalantidium coli: morfología, patogenia y diagnósticoanaNo ratings yet
- Strongyloidiasis: Dr:Jesus Robles Barzola Servicio de Infectologia Hospital Alberto Sabogal SologurenDocument48 pagesStrongyloidiasis: Dr:Jesus Robles Barzola Servicio de Infectologia Hospital Alberto Sabogal SologurenIvone Cruzado UrcoNo ratings yet
- Giardiasis: causas, síntomas y tratamiento de la infección por Giardia lambliaDocument6 pagesGiardiasis: causas, síntomas y tratamiento de la infección por Giardia lambliaSilver Flores100% (1)
- AmebiasisDocument6 pagesAmebiasiseduardoNo ratings yet
- Temas de Segundo Parcial ParacitologiaDocument19 pagesTemas de Segundo Parcial ParacitologiaDario Gutiérrez QuirozNo ratings yet
- Balantidiasis: protozoo causante y caso clínicoDocument10 pagesBalantidiasis: protozoo causante y caso clínicoSandra López JiménezNo ratings yet
- Diapositiva Grupo 7....Document18 pagesDiapositiva Grupo 7....DACFIGER ProductionNo ratings yet
- AMEBIASISDocument7 pagesAMEBIASISRomi Scarlet CanedoNo ratings yet
- Guía de Preguntas Seminario Parasitología: Hystolitica y Cuáles Son Sus Características Más Sobresalientes?Document6 pagesGuía de Preguntas Seminario Parasitología: Hystolitica y Cuáles Son Sus Características Más Sobresalientes?José David Cortez Centeno 20190155No ratings yet
- Resumen - AmebasDocument18 pagesResumen - AmebasIndira PortilloNo ratings yet
- Gastritis Etc y Cancer Estomago PDFDocument229 pagesGastritis Etc y Cancer Estomago PDFNena Guadalupe100% (2)
- Colitis ParasitariaDocument13 pagesColitis ParasitariaSebas MontoyaNo ratings yet
- Helicobacter PiloriDocument9 pagesHelicobacter PiloriLizsavelSVNo ratings yet
- DocumentoDocument25 pagesDocumentoEderth CorderoNo ratings yet
- Balantidium ColiDocument13 pagesBalantidium ColiMaryerlin Gavidia0% (1)
- PrasitosDocument3 pagesPrasitosnayibe monteroNo ratings yet
- Helicobacter PyloriDocument35 pagesHelicobacter PyloriFeraligatrNo ratings yet
- H. pylori gastritis 40Document12 pagesH. pylori gastritis 40yessica Mercado100% (1)
- Infección Por Helicobacter PyloriDocument13 pagesInfección Por Helicobacter PyloriAARON NICOLAS UCHUYPUMA CHOQUECHAMBINo ratings yet
- Parasitosis IntestinalDocument28 pagesParasitosis IntestinalMelany OlivoNo ratings yet
- 16-D-Guevara Castro Evelyn Alexandra-Enfermedades Parasitarias en El NiñoDocument21 pages16-D-Guevara Castro Evelyn Alexandra-Enfermedades Parasitarias en El NiñoEve GuevaraNo ratings yet
- Monografía MODIFICADODocument16 pagesMonografía MODIFICADOdavid jaraNo ratings yet
- Bal Anti Dias IsDocument9 pagesBal Anti Dias IsLeonidas Rafael Guerrero OlivoNo ratings yet
- Helicobacter Pylori y BrucellaDocument7 pagesHelicobacter Pylori y BrucellaDaniel CruzNo ratings yet
- Helicobacter Pylori. DX y ManejoDocument3 pagesHelicobacter Pylori. DX y Manejoairamtoribio100% (1)
- Balantidium Coli ResumenDocument2 pagesBalantidium Coli ResumenMiranda SalinaNo ratings yet
- parasitologia 7-8Document6 pagesparasitologia 7-8David MoraNo ratings yet
- Helycobacter PyloriDocument16 pagesHelycobacter PyloriMar MijangosNo ratings yet
- AmebiasisDocument6 pagesAmebiasisMary Selvy Pereira MonteroNo ratings yet
- PolvosDocument21 pagesPolvosBaiklah MartinezNo ratings yet
- SOLUCIONESDocument3 pagesSOLUCIONESBaiklah MartinezNo ratings yet
- C3Document87 pagesC3Baiklah MartinezNo ratings yet
- ANTIÁCIDOSDocument1 pageANTIÁCIDOSBaiklah MartinezNo ratings yet
- Valores Éticos & Leg. Farmac.Document16 pagesValores Éticos & Leg. Farmac.Baiklah MartinezNo ratings yet
- EucaliptoDocument2 pagesEucaliptoBaiklah MartinezNo ratings yet
- So Luci OnesDocument1 pageSo Luci OnesBaiklah MartinezNo ratings yet
- SOLUCIONESDocument3 pagesSOLUCIONESBaiklah MartinezNo ratings yet
- ANTIÁCIDOSDocument1 pageANTIÁCIDOSBaiklah MartinezNo ratings yet
- JNC 7Document26 pagesJNC 7api-3721195100% (2)
- Introducción a la formulación magistralDocument16 pagesIntroducción a la formulación magistralIvan Camilo Gómez100% (2)
- Carbamazepina MiluDocument11 pagesCarbamazepina MiluBaiklah MartinezNo ratings yet
- Salud para TodosDocument9 pagesSalud para TodosBaiklah MartinezNo ratings yet
- 5.cal BalanzaDocument3 pages5.cal BalanzaBaiklah MartinezNo ratings yet
- 5.cal BalanzaDocument8 pages5.cal BalanzaKari DbsNo ratings yet
- OreganoDocument1 pageOreganoBaiklah MartinezNo ratings yet
- Analisis Fisicoquimico y Microbiologico de Agua para Uso PDFDocument46 pagesAnalisis Fisicoquimico y Microbiologico de Agua para Uso PDFLey Cardona MoralesNo ratings yet
- MiliDocument1 pageMiliBaiklah MartinezNo ratings yet
- 5.cal BalanzaDocument8 pages5.cal BalanzaKari DbsNo ratings yet
- Solicit UdDocument1 pageSolicit UdBaiklah MartinezNo ratings yet
- Inteligencia y placer de saberDocument20 pagesInteligencia y placer de saberMarcelaMateluna0% (1)
- Salud para TodosDocument9 pagesSalud para TodosBaiklah MartinezNo ratings yet
- Acción Del HombreDocument2 pagesAcción Del HombreBaiklah MartinezNo ratings yet
- La industria farmacéutica peruana se internacionalizaDocument8 pagesLa industria farmacéutica peruana se internacionalizaBaiklah MartinezNo ratings yet
- La industria farmacéutica peruana se internacionalizaDocument8 pagesLa industria farmacéutica peruana se internacionalizaBaiklah MartinezNo ratings yet
- 26 Lansoprazol TabletasDocument12 pages26 Lansoprazol TabletasBaiklah MartinezNo ratings yet
- Cro No GramaDocument3 pagesCro No GramaBaiklah MartinezNo ratings yet
- Acción Del HombreDocument2 pagesAcción Del HombreBaiklah MartinezNo ratings yet
- El Proceso Salud EnfermedadDocument10 pagesEl Proceso Salud Enfermedadnutricionobstetricia93% (84)
- Calculo de PrevalenciaDocument19 pagesCalculo de PrevalenciaMarcos CalizayaNo ratings yet
- Formato 2, Correccion Eddy Jose Tubon Torres Rev 23-10-21Document13 pagesFormato 2, Correccion Eddy Jose Tubon Torres Rev 23-10-21Eddy TubonNo ratings yet
- ConclusionesDocument13 pagesConclusionesVictor A. Felipe De la RosaNo ratings yet
- Análisis de tendencias en redes sociales para identificar ideas de negocioDocument18 pagesAnálisis de tendencias en redes sociales para identificar ideas de negocioLuisa Fernandez FlorezNo ratings yet
- Informe Moto LinialDocument29 pagesInforme Moto LinialWilderQuispeRamosNo ratings yet
- Tratamientos para tu finca sana con CalbosDocument36 pagesTratamientos para tu finca sana con CalbosGlenda Martin Rondon100% (3)
- Consentimiento Informado Tattoo P RellenarDocument4 pagesConsentimiento Informado Tattoo P RellenarAudry RamosNo ratings yet
- Alopecia AreataDocument18 pagesAlopecia AreataMauricio LampsonNo ratings yet
- Lipoproteinas 1Document3 pagesLipoproteinas 1Emily PaezNo ratings yet
- Semergen-Tratamiento de La Psoriasis en APDocument48 pagesSemergen-Tratamiento de La Psoriasis en APYamileth LopezNo ratings yet
- Algoritmo diabetesDocument2 pagesAlgoritmo diabetesDavidRodríguezVeigaNo ratings yet
- Segunda PrescripciónDocument17 pagesSegunda PrescripciónYanina TsanganeliasNo ratings yet
- Fo-Gth-71 Formato de Encuesta de Sintomas de Desordenes MusculoesqueleticosDocument1 pageFo-Gth-71 Formato de Encuesta de Sintomas de Desordenes Musculoesqueleticosehiner rodriguez0% (1)
- Indicadores Neo Pi RDocument14 pagesIndicadores Neo Pi RBryam ChaconNo ratings yet
- Neuroproteccion CerebralDocument39 pagesNeuroproteccion Cerebralhipocrates_per100% (1)
- Guía de procedimiento: lipotransferencia facialDocument14 pagesGuía de procedimiento: lipotransferencia facialyithian riveraNo ratings yet
- Funciones y órganos del aparato reproductor femenino bovinoDocument8 pagesFunciones y órganos del aparato reproductor femenino bovinoMilba Soria Bonilla100% (1)
- Nivel conocimiento enfermedad pacientes programa hipertensiónDocument2 pagesNivel conocimiento enfermedad pacientes programa hipertensiónJorge SuarezNo ratings yet
- Seminario Completo Semana 2 - Anemia A Celulas FalciformesDocument12 pagesSeminario Completo Semana 2 - Anemia A Celulas FalciformesKevin ArboledaNo ratings yet
- Introduccion Al Soporte Nutricional.Document24 pagesIntroduccion Al Soporte Nutricional.Paula De La CruzNo ratings yet
- Exportación de paltas Hass a CaliforniaDocument41 pagesExportación de paltas Hass a CaliforniaGiancarlos LajoNo ratings yet
- Plan TBCDocument20 pagesPlan TBCKALIAK54No ratings yet
- Reporte actividades Día Mundial Lucha contra DepresiónDocument1 pageReporte actividades Día Mundial Lucha contra DepresiónJuan HernandezNo ratings yet
- Introducción a la discapacidad visualDocument8 pagesIntroducción a la discapacidad visualDaniel GarcíaNo ratings yet
- EPOCDocument49 pagesEPOCKarla PugaNo ratings yet
- Obesidad PPT FinalDocument58 pagesObesidad PPT FinalMILAGROSNo ratings yet
- Ensayo Clinico Pie PlanoDocument3 pagesEnsayo Clinico Pie PlanoMaribel0% (1)
- Cuatro Signos Cardinales de La Adiccion PDFDocument3 pagesCuatro Signos Cardinales de La Adiccion PDFAmalia García JuradoNo ratings yet
- Antidepresivos Rang & Dale. Farmacología 7º Ed.Document20 pagesAntidepresivos Rang & Dale. Farmacología 7º Ed.Jairo Alexis Ortega RodríguezNo ratings yet
- Síndrome Bronquial ObstructivoDocument5 pagesSíndrome Bronquial ObstructivoConstanza Barrera RiquelmeNo ratings yet
- SEMANA Nro. 01 MICROBIOLOGIA Y PARASITOLOGIA MEDICADocument50 pagesSEMANA Nro. 01 MICROBIOLOGIA Y PARASITOLOGIA MEDICAKaterine Rosaura Blas CernaNo ratings yet