You might also like
- Clonación de Felinos en Peligro Utilizando Ovocitos Donadores Heterospecíficos y Transferencia de Embriones InterespeciesDocument7 pagesClonación de Felinos en Peligro Utilizando Ovocitos Donadores Heterospecíficos y Transferencia de Embriones InterespeciesHeber Rengifo ChotaNo ratings yet
- Conejos Clonados Producidos Por Transferencia Nuclear de Células Somáticas AdultasDocument4 pagesConejos Clonados Producidos Por Transferencia Nuclear de Células Somáticas AdultasHeber Rengifo ChotaNo ratings yet
- Conejos Clonados Producidos Por Transferencia Nuclear de Células Somáticas AdultasDocument4 pagesConejos Clonados Producidos Por Transferencia Nuclear de Células Somáticas AdultasHeber Rengifo ChotaNo ratings yet
- Articulo en Ingles Clonación de La Oveja Dolly PDFDocument5 pagesArticulo en Ingles Clonación de La Oveja Dolly PDFHeber Rengifo ChotaNo ratings yet
- Cerdos Clonados Producidos Por Transferencia Nuclear de Células Somáticas AdultasDocument5 pagesCerdos Clonados Producidos Por Transferencia Nuclear de Células Somáticas AdultasHeber Rengifo ChotaNo ratings yet
- Bipartición EmbrionariaDocument3 pagesBipartición EmbrionariaHeber Rengifo ChotaNo ratings yet
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (890)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Instruction Manual PC Interface RSM 100: TOSHIBA Corporation 1999 All Rights ReservedDocument173 pagesInstruction Manual PC Interface RSM 100: TOSHIBA Corporation 1999 All Rights ReservedFarid HakikiNo ratings yet
- CONTACT DETAILS HC JUDGES LIBRARIESDocument4 pagesCONTACT DETAILS HC JUDGES LIBRARIESSHIVAM BHATTACHARYANo ratings yet
- 2018 JC2 H2 Maths SA2 River Valley High SchoolDocument50 pages2018 JC2 H2 Maths SA2 River Valley High SchoolZtolenstarNo ratings yet
- 4mb1 02r Que 20220608Document32 pages4mb1 02r Que 20220608M.A. HassanNo ratings yet
- Job Interview CV TipsDocument2 pagesJob Interview CV TipsCarlos Moraga Copier100% (1)
- ACCA P1 Governance, Risk, and Ethics - Revision QuestionsDocument2 pagesACCA P1 Governance, Risk, and Ethics - Revision QuestionsChan Tsu ChongNo ratings yet
- 6th GATEDocument33 pages6th GATESamejiel Aseviel LajesielNo ratings yet
- Keb Combivis 6 enDocument232 pagesKeb Combivis 6 enhaithamNo ratings yet
- Going to the cinema listening practiceDocument2 pagesGoing to the cinema listening practiceMichael DÍligo Libre100% (1)
- Guidelines To MAS Notice 626 April 2015Document62 pagesGuidelines To MAS Notice 626 April 2015Wr OngNo ratings yet
- Reg OPSDocument26 pagesReg OPSAlexandru RusuNo ratings yet
- Investigation Report on Engine Room Fire on Ferry BerlinDocument63 pagesInvestigation Report on Engine Room Fire on Ferry Berlin卓文翔No ratings yet
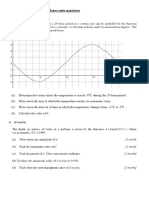
- Sine and Cosine Exam QuestionsDocument8 pagesSine and Cosine Exam QuestionsGamer Shabs100% (1)
- DesignWS P1 PDFDocument673 pagesDesignWS P1 PDFcaubehamchoi6328No ratings yet
- Lease of Residential HouseDocument4 pagesLease of Residential HousedenvergamlosenNo ratings yet
- Sebu6918 03 00 AllDocument94 pagesSebu6918 03 00 AllAhmed Moustafa100% (1)
- AP Research Survival Guide - RevisedDocument58 pagesAP Research Survival Guide - RevisedBadrEddin IsmailNo ratings yet
- The Meaning of Solar CookerDocument4 pagesThe Meaning of Solar CookerJaridah Mat YakobNo ratings yet
- Microscopes Open Up An Entire World That You Can't See With The Naked EyeDocument4 pagesMicroscopes Open Up An Entire World That You Can't See With The Naked EyeLouie Jane EleccionNo ratings yet
- Environmental ScienceDocument5 pagesEnvironmental Sciencearijit_ghosh_18No ratings yet
- Megneto TherapyDocument15 pagesMegneto TherapyedcanalNo ratings yet
- Lehman BrothersDocument10 pagesLehman BrothersJaikishin RuprajNo ratings yet
- CHEMOREMEDIATIONDocument8 pagesCHEMOREMEDIATIONdeltababsNo ratings yet
- Securifire 1000-ExtractedDocument2 pagesSecurifire 1000-ExtractedWilkeey EstrellanesNo ratings yet
- Nidhi Investment Consultant: Magic Mix Illustration For Mr. AB Prafulbhai (Age 18)Document2 pagesNidhi Investment Consultant: Magic Mix Illustration For Mr. AB Prafulbhai (Age 18)jdchandrapal4980No ratings yet
- Nortek Quick Guide: - To Vectrino ProfilerDocument4 pagesNortek Quick Guide: - To Vectrino ProfilerAndresFelipePrietoAlarconNo ratings yet
- Case NoDocument13 pagesCase NoLaurente JessicaNo ratings yet
- Certification of Psychology Specialists Application Form: Cover PageDocument3 pagesCertification of Psychology Specialists Application Form: Cover PageJona Mae MetroNo ratings yet
- C++ Practical FileDocument15 pagesC++ Practical FilePreetish ChandraNo ratings yet
- XLVI ESAN INTERNATIONAL WEEK (MBA Only - July 2023)Document38 pagesXLVI ESAN INTERNATIONAL WEEK (MBA Only - July 2023)Juan Diego Fernández CastilloNo ratings yet