You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- FxbifosfonatoDocument23 pagesFxbifosfonatoAbi HinojosaNo ratings yet
- Hormone Replacement TherapyDocument8 pagesHormone Replacement TherapyWawanNo ratings yet
- SalerioDocument28 pagesSalerioRizqaFebrilianyNo ratings yet
- National Clinical Guidelines For Management of Diabetes Melitus - CompleteDocument73 pagesNational Clinical Guidelines For Management of Diabetes Melitus - CompleteLaurensia Erlina Natalia100% (1)
- Practical Guidelines Blood TransfusionDocument64 pagesPractical Guidelines Blood TransfusionMazen MohamedNo ratings yet
- FSP ORP Handout English Intertrochanteric Femoral Fractures FinalDocument10 pagesFSP ORP Handout English Intertrochanteric Femoral Fractures FinalPratiwi Indah SayektiNo ratings yet
- Vaginaldischarge PDFDocument1 pageVaginaldischarge PDFVidini Kusuma AjiNo ratings yet
- JurnalDocument336 pagesJurnalVidini Kusuma AjiNo ratings yet
- Side-Effects of Topical Steroids: A Long Overdue Revisit: Review ArticleDocument10 pagesSide-Effects of Topical Steroids: A Long Overdue Revisit: Review ArticleVidini Kusuma AjiNo ratings yet
- Transfusion of Human Volunteers With Older, Stored Red Blood Cells Produces Extravascular Hemolysis and Circulating Non-Transferrin-Bound IronDocument9 pagesTransfusion of Human Volunteers With Older, Stored Red Blood Cells Produces Extravascular Hemolysis and Circulating Non-Transferrin-Bound IronVidini Kusuma AjiNo ratings yet
- Wjo 8 242Document15 pagesWjo 8 242Jacob DoughertyNo ratings yet
- WF Lab Clin TM RbcsDocument3 pagesWF Lab Clin TM RbcsVidini Kusuma AjiNo ratings yet
- Kontrasepsi Dan Ket PDFDocument5 pagesKontrasepsi Dan Ket PDFgusriaNo ratings yet
- Acls AlgorithmsDocument8 pagesAcls Algorithmsdydy_7193No ratings yet
- 2017 Article 857Document7 pages2017 Article 857ChangNo ratings yet
- Femur Shaft Fractures in Children: An Epidemiological and Biomechanical StudyDocument71 pagesFemur Shaft Fractures in Children: An Epidemiological and Biomechanical StudyVidini Kusuma AjiNo ratings yet
- WF Lab Clin TM RbcsDocument3 pagesWF Lab Clin TM RbcsVidini Kusuma AjiNo ratings yet
- JurnalDocument7 pagesJurnalVidini Kusuma AjiNo ratings yet
- JurnalDocument4 pagesJurnalVidini Kusuma AjiNo ratings yet
- Mueller Ao Class PDFDocument10 pagesMueller Ao Class PDFTrandafir LacramioaraNo ratings yet
- 2017 GINA Report, Global Strategy For Asthma Management and Prevention PDFDocument159 pages2017 GINA Report, Global Strategy For Asthma Management and Prevention PDFNalemi JT100% (1)
- Copd PDFDocument44 pagesCopd PDFSusiYanuariNo ratings yet
- Role of AdjuvantDocument5 pagesRole of AdjuvantMutiaraNo ratings yet
- The Revised 2014 GINA Strategy Report: Opportunities For ChangeDocument7 pagesThe Revised 2014 GINA Strategy Report: Opportunities For ChangeVidini Kusuma AjiNo ratings yet
- 5 ArticleDocument6 pages5 ArticleVidini Kusuma AjiNo ratings yet
- PaperDocument4 pagesPaperVidini Kusuma AjiNo ratings yet
- Correlation Between Family Health Task and Relapse of SchizophreniaDocument14 pagesCorrelation Between Family Health Task and Relapse of SchizophreniaFitri GessiaNo ratings yet
- Jurnal THTDocument5 pagesJurnal THTSandra MignonNo ratings yet
- 2015 Ann R Coll Surg Engl Management of Open Lower Limb Injuries in South West England and WalesDocument5 pages2015 Ann R Coll Surg Engl Management of Open Lower Limb Injuries in South West England and WalesVidini Kusuma AjiNo ratings yet
- Vol.18 No.3 6Document73 pagesVol.18 No.3 6Vidini Kusuma AjiNo ratings yet
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (894)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- 2 Part 1Document99 pages2 Part 1m_luchianNo ratings yet
- Citric Acid Production Stirred TankDocument9 pagesCitric Acid Production Stirred TankKarliiux MedinaNo ratings yet
- Chemical Composition of Meat and Nut Water of Major Coconut in PakistanDocument7 pagesChemical Composition of Meat and Nut Water of Major Coconut in PakistanSkyle VoNo ratings yet
- Theories of Tooth MovementDocument11 pagesTheories of Tooth MovementAhmedsy Ahmedsy AhmedsyNo ratings yet
- Pengantar Mikrobiologi LingkunganDocument28 pagesPengantar Mikrobiologi LingkunganAhmad SyahrulNo ratings yet
- Molecular Genetics of Colorectal Cancer - UpToDateDocument41 pagesMolecular Genetics of Colorectal Cancer - UpToDateToweran ToweraneNo ratings yet
- Candida Tropicalis - ACMGDocument12 pagesCandida Tropicalis - ACMGSergio Iván López LallanaNo ratings yet
- 03 Chapter 3Document148 pages03 Chapter 3arghaNo ratings yet
- Full Download Human Physiology From Cells To Systems 8th Edition Lauralee Sherwood Solutions ManualDocument36 pagesFull Download Human Physiology From Cells To Systems 8th Edition Lauralee Sherwood Solutions Manualgambolrapinous.ggqcdr100% (39)
- Turbidimetry and Nephelometry1Document3 pagesTurbidimetry and Nephelometry1maxim_crank6101100% (1)
- Ssac 2104Document100 pagesSsac 2104Amrutha amrNo ratings yet
- Forest Health and Biotechnology - Possibilities and Considerations (2019)Document241 pagesForest Health and Biotechnology - Possibilities and Considerations (2019)apertusNo ratings yet
- Module 2 NCM 114Document11 pagesModule 2 NCM 114Erven AranasNo ratings yet
- Chapter 2 Medical Terminology Verified AnswersDocument5 pagesChapter 2 Medical Terminology Verified AnswersGregg ProducerNo ratings yet
- Biology Investigatory ProjectDocument17 pagesBiology Investigatory ProjectAnmol Dhungel100% (6)
- Pemphigus Subtypes Clinical Features Diagnosis andDocument23 pagesPemphigus Subtypes Clinical Features Diagnosis andAnonymous bdFllrgorzNo ratings yet
- Trunking CatalogDocument8 pagesTrunking CatalogDenuka PathiranaNo ratings yet
- Protochordata FIXDocument33 pagesProtochordata FIXSylvia AnggraeniNo ratings yet
- Biomimicry Avjeet SinghDocument21 pagesBiomimicry Avjeet SinghAvjeet SinghNo ratings yet
- Significance of 4D Printing For Dentistry: Materials, Process, and PotentialsDocument9 pagesSignificance of 4D Printing For Dentistry: Materials, Process, and PotentialsMohamed FaizalNo ratings yet
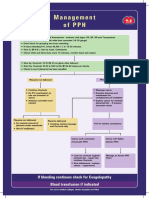
- Management of PPHDocument1 pageManagement of PPH098 U.KARTHIK SARAVANA KANTHNo ratings yet
- IN SEARCH OF SEXUAL ENHANCEMENT THROUGH PILLS AND DRUGSDocument16 pagesIN SEARCH OF SEXUAL ENHANCEMENT THROUGH PILLS AND DRUGSJakob AndradeNo ratings yet
- SOP EcoliDocument3 pagesSOP EcoliIeqa HaziqahNo ratings yet
- UAS Writing For Discourse - 4 Pagi OkDocument1 pageUAS Writing For Discourse - 4 Pagi Oksalwa syabinaNo ratings yet
- Environmental Science: GE 15 - Subject Code: 1603Document18 pagesEnvironmental Science: GE 15 - Subject Code: 1603japheth louie m. gofredoNo ratings yet
- Gazzaniga The Ethical - Brain.the - Science.of - Our.moral - DilemmasDocument229 pagesGazzaniga The Ethical - Brain.the - Science.of - Our.moral - Dilemmaskid_latigo100% (9)
- The Vulcanization of BrainDocument30 pagesThe Vulcanization of BrainJohnny FragosoNo ratings yet
- NSABB Final Report Recommendations Evaluation Oversight Proposed Gain of Function Research May 2016Document109 pagesNSABB Final Report Recommendations Evaluation Oversight Proposed Gain of Function Research May 2016Jim Hoft100% (1)
- Reproductive Biology and Phylogeny of Birds PDFDocument624 pagesReproductive Biology and Phylogeny of Birds PDFFabricio GZNo ratings yet
- 皮膚病介紹Document93 pages皮膚病介紹MK CameraNo ratings yet