You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (120)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Efedrin JurnalDocument4 pagesEfedrin JurnaldesyNo ratings yet
- Heryani, Maryani Akar Kuning Kandungan Aja PDFDocument7 pagesHeryani, Maryani Akar Kuning Kandungan Aja PDFdesyNo ratings yet
- Jurnal Mikro 1 HariDocument6 pagesJurnal Mikro 1 HaridesyNo ratings yet
- Parasit JurnalDocument10 pagesParasit JurnaldesyNo ratings yet
- Intracellular Obligat - Chlamydia PDFDocument45 pagesIntracellular Obligat - Chlamydia PDFdesyNo ratings yet
- Intracellular Obligat - ChlamydiaDocument17 pagesIntracellular Obligat - ChlamydiadesyNo ratings yet
- ChlamydiaDocument45 pagesChlamydiamnn164No ratings yet
- Trauma Medula SpinalisDocument54 pagesTrauma Medula SpinalisdesyNo ratings yet
- 'VIRUS HEPATITIS (Aceri3-PC's Conflicted Copy 2018-03-25) .PPT'Document60 pages'VIRUS HEPATITIS (Aceri3-PC's Conflicted Copy 2018-03-25) .PPT'desyNo ratings yet
- Juhairina: Medical Nutrition Therapy For Disorders of The Lower Gastrointestinal TractDocument82 pagesJuhairina: Medical Nutrition Therapy For Disorders of The Lower Gastrointestinal Tractdesy100% (1)
- Curriculum Vitae: Medical Student Goes Abroad 2018Document3 pagesCurriculum Vitae: Medical Student Goes Abroad 2018desyNo ratings yet
- An Tropo MetriDocument29 pagesAn Tropo MetridesyNo ratings yet
- Sepsis: Nani ZaitunDocument55 pagesSepsis: Nani ZaitundesyNo ratings yet
- Intracellular Obligat - ChlamydiaDocument2 pagesIntracellular Obligat - ChlamydiadesyNo ratings yet
- Jamur Penyebab Infeksi Respirasi PDFDocument122 pagesJamur Penyebab Infeksi Respirasi PDFdesyNo ratings yet
- Intracellular Obligat - ChlamydiaDocument17 pagesIntracellular Obligat - ChlamydiadesyNo ratings yet
- Acute Respiratory Distress Syndrome (ARDS)Document72 pagesAcute Respiratory Distress Syndrome (ARDS)desyNo ratings yet
- Intracellular Obligat - Ricketsia FamilyDocument76 pagesIntracellular Obligat - Ricketsia FamilydesyNo ratings yet
- Nutrisi Pada Penyakit KVDocument66 pagesNutrisi Pada Penyakit KVdesyNo ratings yet
- DM KadDocument18 pagesDM KadWahyu SandikaNo ratings yet
- Endocrine System Pathology 2009Document195 pagesEndocrine System Pathology 2009SofiaemanginteNo ratings yet
- Nutrition For Gout: by JuhairinaDocument39 pagesNutrition For Gout: by Juhairinadesy100% (1)
- ChlamydiaDocument45 pagesChlamydiamnn164No ratings yet
- ChlamydiaDocument45 pagesChlamydiamnn164No ratings yet
- Anatomi Blok 1.5 Brain & Its Surrounding StructuresDocument8 pagesAnatomi Blok 1.5 Brain & Its Surrounding StructuresSabila RosyidaNo ratings yet
- He Ma To PoiesisDocument121 pagesHe Ma To PoiesisdesyNo ratings yet
- 458 4194 1 PBDocument8 pages458 4194 1 PBdesyNo ratings yet
- Conduction System and Prenatal CirculationDocument1 pageConduction System and Prenatal CirculationdesyNo ratings yet
- 458 4194 1 PBDocument8 pages458 4194 1 PBdesyNo ratings yet
- Wollo UniversityDocument14 pagesWollo UniversityabdishakurNo ratings yet
- Perineal CareDocument10 pagesPerineal Careoxidalaj100% (11)
- Laparoscopsic Insemination of Sheep in Republic of Serbia - A.Milovanović, N. Maksimović, T. Barna, M. Lazarević, N. DelićDocument8 pagesLaparoscopsic Insemination of Sheep in Republic of Serbia - A.Milovanović, N. Maksimović, T. Barna, M. Lazarević, N. DelićCk_psihNo ratings yet
- MeiosisDocument42 pagesMeiosisSindhu BabuNo ratings yet
- Nursing Process - Placenta PreviaDocument56 pagesNursing Process - Placenta PreviaRicardo Robianes BautistaNo ratings yet
- Severe Birth Asphyxia: Risk Factors As Seen in A Tertiary Institution in The Niger Delta Area of NigeriaDocument9 pagesSevere Birth Asphyxia: Risk Factors As Seen in A Tertiary Institution in The Niger Delta Area of NigeriaSteven JonesNo ratings yet
- Cudurada Kudhaca XoolahaDocument11 pagesCudurada Kudhaca XoolahaAnonymous ZeRmTfJY83% (12)
- Anatomy & Physiology: The Endocrine SystemDocument1 pageAnatomy & Physiology: The Endocrine SystemEliezer NuenayNo ratings yet
- Assessment of Condom Use For Prevention of Hiv/Aids Among Jimma University Students, Jimmatown, South West EthiopiaDocument42 pagesAssessment of Condom Use For Prevention of Hiv/Aids Among Jimma University Students, Jimmatown, South West EthiopiaFitalaBokaNo ratings yet
- Fascio LaDocument20 pagesFascio LaMuhammad NoorNo ratings yet
- Female Reproductive SystemDocument7 pagesFemale Reproductive SystemRem M RollinasNo ratings yet
- Workbook On Science Science: (Grade 5)Document163 pagesWorkbook On Science Science: (Grade 5)Che LV75% (4)
- Plant Kingdom Previous Year QuestionsDocument4 pagesPlant Kingdom Previous Year QuestionsNavas evNo ratings yet
- Carroll Chapter 05.female AnatomyDocument50 pagesCarroll Chapter 05.female AnatomySitaramNo ratings yet
- Spearmint Tea For PCOSDocument5 pagesSpearmint Tea For PCOSFeeroza KhanNo ratings yet
- International RRLDocument4 pagesInternational RRLXela Asyz SiloNo ratings yet
- Nocturnal Emissions: "Wet Dreams"Document10 pagesNocturnal Emissions: "Wet Dreams"Hanako Shiga100% (1)
- Bsed Fil 1101 - Ed101 - Module 11 - Group 5Document19 pagesBsed Fil 1101 - Ed101 - Module 11 - Group 5Jessabelle D CasanovaNo ratings yet
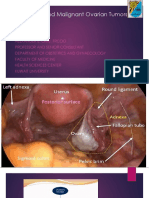
- (#5) Benign and Malignant Ovariaan TumorsDocument35 pages(#5) Benign and Malignant Ovariaan Tumorsmarina_shawkyNo ratings yet
- LactationDocument24 pagesLactationJenyl Bajado100% (1)
- Lyiny inDocument11 pagesLyiny inSi Kaye100% (3)
- стендге тем план гинекология 5курсDocument3 pagesстендге тем план гинекология 5курсНургулNo ratings yet
- STUDENT Tasks For NCM Culminating ProgramDocument4 pagesSTUDENT Tasks For NCM Culminating ProgramLyra LorcaNo ratings yet
- Latihan Kelas9Document12 pagesLatihan Kelas9Afrijal DahrinNo ratings yet
- CC, HPV & HPV VaccineDocument16 pagesCC, HPV & HPV VaccineBalqis BaslemanNo ratings yet
- Nursing Questions& AnswersDocument37 pagesNursing Questions& AnswersSanjeev KumarNo ratings yet
- A Sporadic Case of Delayed Implantation After In-Vitro Fertilization in The Human?Document4 pagesA Sporadic Case of Delayed Implantation After In-Vitro Fertilization in The Human?gardener10No ratings yet
- Eshre Revised Guidelines For Good Practice in IVF LaboratoriesDocument10 pagesEshre Revised Guidelines For Good Practice in IVF LaboratoriesWael BadranNo ratings yet
- Rubia Cordifolia Linn.Document4 pagesRubia Cordifolia Linn.majiamal99No ratings yet
- Mitosis and MeiosisDocument8 pagesMitosis and Meiosisabisantiago6131No ratings yet