You might also like
- Caballo PDFDocument25 pagesCaballo PDFMaría BelénNo ratings yet
- Lucía Willenpart El Tango: Temas y MotivosDocument12 pagesLucía Willenpart El Tango: Temas y MotivosCamilo100% (2)
- ReglasDocument1 pageReglasMaría BelénNo ratings yet
- Bolillero y Cronograma de Bioquímica de La NutriciónDocument7 pagesBolillero y Cronograma de Bioquímica de La NutriciónMaría BelénNo ratings yet
- Reglas Sheriff de NottinghamDocument8 pagesReglas Sheriff de NottinghamCesar Antonio50% (2)
- Cic in RespuestasDocument1 pageCic in RespuestasMaría BelénNo ratings yet
- Pardo y PolarDocument12 pagesPardo y PolarMaría BelénNo ratings yet
- Mateo - Armando DiscépoloDocument10 pagesMateo - Armando DiscépoloGonzalo Marin67% (6)
- Ficha Del AlumnoDocument2 pagesFicha Del AlumnoMaría BelénNo ratings yet
- ANESTÉSIC..Document13 pagesANESTÉSIC..María BelénNo ratings yet
- Cic in Web Contenidos 2015Document8 pagesCic in Web Contenidos 2015CeciliaNo ratings yet
- Jorge Luis Borges - El SurDocument5 pagesJorge Luis Borges - El Suranon_597941881No ratings yet
- 1 SMDocument7 pages1 SMMaría BelénNo ratings yet
- Mijail BajtinDocument8 pagesMijail BajtinNora Natalia LerenaNo ratings yet
- Cronograma Juana y ValentinaDocument1 pageCronograma Juana y ValentinaMaría BelénNo ratings yet
- Literatura y Cine (Crespo)Document9 pagesLiteratura y Cine (Crespo)lorenaNo ratings yet
- Hipouricemiantes, Antigotosos y GlucocorticoidesDocument7 pagesHipouricemiantes, Antigotosos y GlucocorticoidesMaría BelénNo ratings yet
- Farmacología ClínicaDocument4 pagesFarmacología ClínicaMaría BelénNo ratings yet
- Metabolismo Fosfocálcico - Hormonas Sexuales2Document6 pagesMetabolismo Fosfocálcico - Hormonas Sexuales2Brenda TorresNo ratings yet
- Reglas Sheriff de NottinghamDocument8 pagesReglas Sheriff de NottinghamCesar Antonio50% (2)
- Repaso NeurotransmisiónDocument2 pagesRepaso NeurotransmisiónMaría BelénNo ratings yet
- Neurotransmisión AdrenérgicaDocument6 pagesNeurotransmisión AdrenérgicaMaría BelénNo ratings yet
- Farmacodinamia IDocument4 pagesFarmacodinamia IBrenda TorresNo ratings yet
- Personaje 1Document1 pagePersonaje 1María BelénNo ratings yet
- Farmacocinética: Absorción, Distribución y MetabolismoDocument5 pagesFarmacocinética: Absorción, Distribución y MetabolismoBenjamín GonzálezNo ratings yet
- Neurotransmisión AdrenérgicaDocument6 pagesNeurotransmisión AdrenérgicaMaría BelénNo ratings yet
- Reglas Sheriff de NottinghamDocument8 pagesReglas Sheriff de NottinghamCesar Antonio50% (2)
- Clase 2 de Vocabulario Unidades 5 y 6Document6 pagesClase 2 de Vocabulario Unidades 5 y 6María BelénNo ratings yet
- Clase 1 de Vocabulario Unidades 1,2 Y3Document9 pagesClase 1 de Vocabulario Unidades 1,2 Y3María BelénNo ratings yet
- 3.1.C Acento DiacríticoDocument4 pages3.1.C Acento DiacríticoSelma AlbuquerqueNo ratings yet
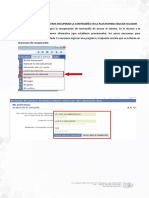
- Manual Recuperar Contraseña de La Plataforma EducarecuadorDocument6 pagesManual Recuperar Contraseña de La Plataforma Educarecuadorrobertotenezaca100% (1)
- Presentación de Costos de OperaciónDocument27 pagesPresentación de Costos de Operaciónmariogut2000No ratings yet
- Tarea Salario Minimo.Document8 pagesTarea Salario Minimo.Nebin PeñaNo ratings yet
- RVM #049-2022-Minedu PDFDocument117 pagesRVM #049-2022-Minedu PDFFran Dux Soncco PuchoNo ratings yet
- Historia de Poza RicaDocument1 pageHistoria de Poza RicaAtzhiri VenturaNo ratings yet
- MP110 Instalador Central de Alarma Manual de InstalaciónDocument68 pagesMP110 Instalador Central de Alarma Manual de InstalaciónJesus GarciaNo ratings yet
- T 2317 IndDocument248 pagesT 2317 IndWashington LendroNo ratings yet
- AATCC Test Method 61 2009.en - EsDocument5 pagesAATCC Test Method 61 2009.en - EsJhon EdissonNo ratings yet
- Tomar el último lugarDocument4 pagesTomar el último lugarGabriel Alonso VazquezNo ratings yet
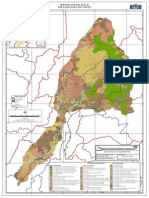
- Rio Vinces Uso Del SueloDocument1 pageRio Vinces Uso Del SuelocrematorioNo ratings yet
- Hoja 1 Constitución Política de GuatemalaDocument5 pagesHoja 1 Constitución Política de GuatemalaJeannette Fuentes DiazNo ratings yet
- Qué Se Necesita Saber para Tener Éxito en Los Hechizos para EnamorarDocument2 pagesQué Se Necesita Saber para Tener Éxito en Los Hechizos para EnamorarhechizosNo ratings yet
- Municipalidad de MiracostaDocument2 pagesMunicipalidad de MiracostaJian SotoNo ratings yet
- El Hombre Que Deseaba OroDocument5 pagesEl Hombre Que Deseaba OroEliana CogolloNo ratings yet
- La civilización europea y el mundo islámicoDocument11 pagesLa civilización europea y el mundo islámicoLira ValentinaNo ratings yet
- Visual Basic 6 0 UltimoDocument65 pagesVisual Basic 6 0 UltimoWILSONNo ratings yet
- Guía Género Lírico EvaluadaDocument4 pagesGuía Género Lírico EvaluadaClaudia Andrea Mera VargasNo ratings yet
- Reglamento BHSTV 2022Document9 pagesReglamento BHSTV 2022Fernanda AguilarNo ratings yet
- Análisis Comparativo de Cuadros de PreciosDocument8 pagesAnálisis Comparativo de Cuadros de PreciosGerardo GarcíaNo ratings yet
- La Responsabilidad y Etica Profesional MVZ - LeyesDocument81 pagesLa Responsabilidad y Etica Profesional MVZ - LeyesLaura J Guerrero100% (2)
- Informaciongeneraldelaasignatura PDFDocument18 pagesInformaciongeneraldelaasignatura PDFangel2evangelisNo ratings yet
- Guía de Aprendizajes LENG Cuarto Básico Semana 29 de AgostoDocument2 pagesGuía de Aprendizajes LENG Cuarto Básico Semana 29 de AgostohaylinakjdNo ratings yet
- Tipos de Documentos AdministrativosDocument44 pagesTipos de Documentos AdministrativosPiva Binlex100% (2)
- Qué Es El RelativismoDocument5 pagesQué Es El RelativismoBrunoMioNo ratings yet
- Backbone Nacional PDFDocument40 pagesBackbone Nacional PDFmariusNo ratings yet
- IM2 - Resumen - Paola Vidal (2015) PDFDocument60 pagesIM2 - Resumen - Paola Vidal (2015) PDFRoberto Ignacio PobleteNo ratings yet
- Actividad de Matemáticas 5 de Matemáticas - Grado 10-Yalidis v-2021Document2 pagesActividad de Matemáticas 5 de Matemáticas - Grado 10-Yalidis v-2021Luis Fernando Cogollo CavadiaNo ratings yet
- Solidworks Composer CWDocument4 pagesSolidworks Composer CWJosephRomeraloNo ratings yet
- Caracteristicas Generales Del Derecho AnglosajonDocument3 pagesCaracteristicas Generales Del Derecho AnglosajonLuis Augusto Zenteno Enriquez100% (1)