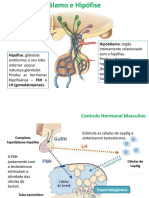
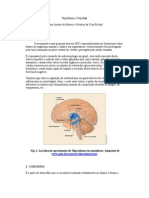
You might also like
- Controle Neuroendócrino Da ReproduçãoDocument27 pagesControle Neuroendócrino Da ReproduçãoLuiz Henrique Ferreira100% (2)
- 995 - Aula Do Eixo Hipotalamico Hipofisario Gonadal UterinoDocument57 pages995 - Aula Do Eixo Hipotalamico Hipofisario Gonadal UterinoJoão Paulo Da MataNo ratings yet
- ACFrOgCaXJQNEG9Owg1r 1nBdf3paTpn6PTuwxhmTe9pn EhYKwzr - CqZsASOav0CcgVrWGf45vFKnCFqpIuRSFowwOeVQwVF1cvYrt9rO9ezjxgYj2Q UKX3lKiJctEnWnvi5gK9uFh0S16JLUDocument21 pagesACFrOgCaXJQNEG9Owg1r 1nBdf3paTpn6PTuwxhmTe9pn EhYKwzr - CqZsASOav0CcgVrWGf45vFKnCFqpIuRSFowwOeVQwVF1cvYrt9rO9ezjxgYj2Q UKX3lKiJctEnWnvi5gK9uFh0S16JLULucca Dal MoroNo ratings yet
- Anticoncepcionais Hormonais Mari QuintelaDocument8 pagesAnticoncepcionais Hormonais Mari QuintelaAfiliado LendárioNo ratings yet
- Fisiologia Do Ciclo Menstrual Normal (Dra Karin)Document5 pagesFisiologia Do Ciclo Menstrual Normal (Dra Karin)Laura BrustolinNo ratings yet
- Ciclo MenstrualDocument3 pagesCiclo MenstrualThaiza LaudaresNo ratings yet
- Ciclo Menstrual Unama AtualDocument59 pagesCiclo Menstrual Unama AtualEly Paula de OliveiraNo ratings yet
- Nascimento PROBLEMA 08Document13 pagesNascimento PROBLEMA 08João CassorieloNo ratings yet
- Ginecolog (1)Document65 pagesGinecolog (1)Luiza Lopes MarouelliNo ratings yet
- Regulação Hormonal MasculinaDocument2 pagesRegulação Hormonal MasculinaMarta OliveiraNo ratings yet
- Reprodução - Fisiologia e Endocrinologia Do Ciclo EstralDocument13 pagesReprodução - Fisiologia e Endocrinologia Do Ciclo EstralRebeca MoreiraNo ratings yet
- 9 Fisiologia EndócrinaDocument13 pages9 Fisiologia EndócrinaThiago Henrique Oliveira100% (1)
- Fisiologia Do Ciclo MenstrualDocument7 pagesFisiologia Do Ciclo Menstrualbarbara.dourado12484No ratings yet
- (EMBRIOLOGIA - RESUMO) Controle Hormonal Da Ovogênese e Da EspermatogêneseDocument6 pages(EMBRIOLOGIA - RESUMO) Controle Hormonal Da Ovogênese e Da EspermatogêneseCaroline Milheiro da SilvaNo ratings yet
- 2 - Regulacao Hormonal Homem e MulherDocument10 pages2 - Regulacao Hormonal Homem e MulherCarina GasparNo ratings yet
- Ciclo MenstrualDocument11 pagesCiclo MenstrualNaro SantNo ratings yet
- Ciclo MenstrualDocument9 pagesCiclo Menstrualraquel jepsenNo ratings yet
- Eixo Hipotálamo-PituitáriaDocument2 pagesEixo Hipotálamo-PituitáriaCoconut TcmNo ratings yet
- Resumed - SUADocument7 pagesResumed - SUAIsabel GomesNo ratings yet
- Apostila Base Da Medicina 2022 - Ginecologia (1) - 35-59Document25 pagesApostila Base Da Medicina 2022 - Ginecologia (1) - 35-59Larissa MerciellyNo ratings yet
- A Interpretaçoes de ResultadosDocument1 pageA Interpretaçoes de ResultadosDenise BengeNo ratings yet
- SP1 - Sem Lenco Sem DocumentoDocument8 pagesSP1 - Sem Lenco Sem DocumentoRenan VillacaNo ratings yet
- Fisiologia Do Ciclo MenstrualDocument23 pagesFisiologia Do Ciclo MenstrualKarittaNo ratings yet
- Fisiologia Módulo 8 (Endócrino)Document18 pagesFisiologia Módulo 8 (Endócrino)vargas.pedro2003No ratings yet
- Síndrome Dos Ovários PolicísticosDocument2 pagesSíndrome Dos Ovários PolicísticosMaria Helena MartinsNo ratings yet
- Endocrinologia Da ReproduçãoDocument88 pagesEndocrinologia Da ReproduçãoJuan HernandezNo ratings yet
- Aula 4 - Regulação Reprodução Fêmea e SazonalidadeDocument19 pagesAula 4 - Regulação Reprodução Fêmea e SazonalidadePaloma CraNo ratings yet
- Fisiologia Do Sistema EndócrinoDocument7 pagesFisiologia Do Sistema Endócrinoargosalves3091No ratings yet
- Ginecologia Endócrina - Febrasgo 2010Document173 pagesGinecologia Endócrina - Febrasgo 2010Marcella GarciaNo ratings yet
- Biologia 12º AnoDocument13 pagesBiologia 12º AnoBeatrizNo ratings yet
- Resumo - Video Cilco MenstrualDocument2 pagesResumo - Video Cilco MenstrualVinicio CoelhoNo ratings yet
- Fisiologia IIIDocument5 pagesFisiologia IIIAlisson FelipeNo ratings yet
- REPRODUÇÃO em Medicina VeterináriaDocument54 pagesREPRODUÇÃO em Medicina Veterináriajamilesada100% (11)
- Sistema EndócrinoDocument28 pagesSistema EndócrinoBruno Felipe100% (1)
- Problema 2 - Tutoria - Ciclo Menstrual e ClimatérioDocument10 pagesProblema 2 - Tutoria - Ciclo Menstrual e ClimatérioYasmin NascimentoNo ratings yet
- REPRODUÇÃO NeuroendócrinoDocument27 pagesREPRODUÇÃO NeuroendócrinoLarissa AndradeNo ratings yet
- (RETARDO PUBERAL) Lúcio Vilar - Endocrinologia Clínica (2021, Editora Guanabara Koogan) - Libgen - Li - ExtractedDocument20 pages(RETARDO PUBERAL) Lúcio Vilar - Endocrinologia Clínica (2021, Editora Guanabara Koogan) - Libgen - Li - Extractedanna.vieiraNo ratings yet
- Fixação REPRODUÇÃO NeuroendócrinoDocument30 pagesFixação REPRODUÇÃO NeuroendócrinoLarissa AndradeNo ratings yet
- Farmacologia Do S. Reprodutor MasculinoDocument8 pagesFarmacologia Do S. Reprodutor MasculinoLuanna MotterNo ratings yet
- Fisiologia - Eixo Hipotálamo-HipofisárioDocument3 pagesFisiologia - Eixo Hipotálamo-HipofisárioEduvaldo JúniorNo ratings yet
- Trabalho - Hormonas Hipotálamo HipófiseDocument44 pagesTrabalho - Hormonas Hipotálamo Hipófiseturma4md100% (2)
- Sistema Talâmico HipofisárioDocument7 pagesSistema Talâmico HipofisárioGiovanna MazzettiNo ratings yet
- A3 - Sistema Digestório e EndócrinoDocument4 pagesA3 - Sistema Digestório e EndócrinoLuciana100% (1)
- Farmacologia Dos Hormônios SexuaisDocument15 pagesFarmacologia Dos Hormônios SexuaisIzabela SilvaNo ratings yet
- Estudo Dirigido de Fisiologia Endócrina Luciana Gentile GabaritoDocument7 pagesEstudo Dirigido de Fisiologia Endócrina Luciana Gentile GabaritoWellington SilvaNo ratings yet
- Hipotálamo e HipófiseDocument11 pagesHipotálamo e HipófiseVet_arquivos100% (2)
- Pr2 - Saúde Da Mulher 02Document19 pagesPr2 - Saúde Da Mulher 02Luana CristinaNo ratings yet
- Homônios Da Reprodução - ManéDocument3 pagesHomônios Da Reprodução - ManéClínica Veterinária AnimáliaNo ratings yet
- Regulação Hormonal Do Ciclo Sexual FemeninoDocument2 pagesRegulação Hormonal Do Ciclo Sexual FemeninoMarta OliveiraNo ratings yet
- Hormonioterapia em Pequenos AnimaisDocument18 pagesHormonioterapia em Pequenos AnimaisAline Schmidt100% (1)
- Fisiolgia Humana 8Document6 pagesFisiolgia Humana 8Alessandro PedrettiNo ratings yet
- Capítulo 80 - Funções Reprodutivas e Hormonais Masculinas (E Função Da Glândula Pineal) - 1 PáginaDocument1 pageCapítulo 80 - Funções Reprodutivas e Hormonais Masculinas (E Função Da Glândula Pineal) - 1 PáginaandreamokadaNo ratings yet
- Resumo ReproduçãoDocument5 pagesResumo ReproduçãoBe MottaNo ratings yet
- RESUMO Gineco ObstDocument80 pagesRESUMO Gineco ObstDiego Desconci100% (2)
- A Infertilidade Não É Apenas Um Problema Do CasalFrom EverandA Infertilidade Não É Apenas Um Problema Do CasalRating: 2 out of 5 stars2/5 (1)
- Prolactina e Diabetes Melito do tipo 2: o efeito protetor de um hormônio sobre o metabolismo glicídicoFrom EverandProlactina e Diabetes Melito do tipo 2: o efeito protetor de um hormônio sobre o metabolismo glicídicoNo ratings yet
- Equilíbrio hormonal _ Recupere equilíbrio hormonal, libido, sono e emagreça já!From EverandEquilíbrio hormonal _ Recupere equilíbrio hormonal, libido, sono e emagreça já!Rating: 5 out of 5 stars5/5 (2)
- Conteudo ProgramaticcoDocument3 pagesConteudo ProgramaticcoAline Moura Savassini LucenaNo ratings yet
- Inspeção e Tecnologia de Produtos de Origem Animal IIDocument31 pagesInspeção e Tecnologia de Produtos de Origem Animal IIAline Moura Savassini LucenaNo ratings yet
- 4.3 Clinica Medica e Cirurgica de Grandes AnimaisDocument77 pages4.3 Clinica Medica e Cirurgica de Grandes AnimaisAline Moura Savassini LucenaNo ratings yet
- ManualGTACaprinoseOvinos5 0Document13 pagesManualGTACaprinoseOvinos5 0Aline Moura Savassini LucenaNo ratings yet
- Inspeção 01Document44 pagesInspeção 01Rafaela CamposNo ratings yet
- Top!!! PDFDocument23 pagesTop!!! PDFAline Moura Savassini LucenaNo ratings yet
- Manual de Preenchimento para Emissão de Guia de Trânsito Animal de Abelhas, Bicho-Da-Seda E Outros Invertebrados TerrestresDocument6 pagesManual de Preenchimento para Emissão de Guia de Trânsito Animal de Abelhas, Bicho-Da-Seda E Outros Invertebrados TerrestresAline Moura Savassini LucenaNo ratings yet
- Inspeção e Tecnologia de Produtos de Origem Animal IIDocument39 pagesInspeção e Tecnologia de Produtos de Origem Animal IIAline Moura Savassini Lucena100% (2)
- Informática para Concursos Linux PDFDocument41 pagesInformática para Concursos Linux PDFMarcos César100% (1)
- Outlook 2007Document34 pagesOutlook 2007Aline Moura Savassini LucenaNo ratings yet
- Resumo INSS Informática1 PDFDocument16 pagesResumo INSS Informática1 PDFFrancisco Eronilson100% (1)
- WindowsDocument30 pagesWindowsAline Moura Savassini LucenaNo ratings yet
- Noções de Sistema Operacional - Windows e Linux - Informática para Concursos - Ok ConcursosDocument6 pagesNoções de Sistema Operacional - Windows e Linux - Informática para Concursos - Ok ConcursosAline Moura Savassini LucenaNo ratings yet
- BackupDocument18 pagesBackupAline Moura Savassini LucenaNo ratings yet
- Navegador de Internet e Correio Eletrônico - Informática para Concursos - Ok ConcursosDocument2 pagesNavegador de Internet e Correio Eletrônico - Informática para Concursos - Ok ConcursosAline Moura Savassini LucenaNo ratings yet
- Engenharia de Software - Informática para Concursos - Apostila Grátis - Ok ConcursosDocument3 pagesEngenharia de Software - Informática para Concursos - Apostila Grátis - Ok ConcursosAline Moura Savassini LucenaNo ratings yet
- Navegador de Internet e Correio Eletrônico - Informática para Concursos - Ok ConcursosDocument3 pagesNavegador de Internet e Correio Eletrônico - Informática para Concursos - Ok ConcursosAline Moura Savassini LucenaNo ratings yet
- Noções de Sistema Operacional - Windows e Linux - Informática para Concursos - Ok ConcursosDocument3 pagesNoções de Sistema Operacional - Windows e Linux - Informática para Concursos - Ok ConcursosAline Moura Savassini LucenaNo ratings yet
- Linguagem de Programação - Informática para Concursos - Apostila Grátis - Ok ConcursosDocument3 pagesLinguagem de Programação - Informática para Concursos - Apostila Grátis - Ok ConcursosAline Moura Savassini LucenaNo ratings yet
- Engenharia de Software - Informática para Concursos - Apostila Grátis - Ok ConcursosDocument3 pagesEngenharia de Software - Informática para Concursos - Apostila Grátis - Ok ConcursosAline Moura Savassini LucenaNo ratings yet
- Equipe CirurgicaDocument19 pagesEquipe CirurgicaAline Moura Savassini LucenaNo ratings yet
- Navegador de Internet e Correio Eletrônico - Informática para Concursos - Ok ConcursosDocument2 pagesNavegador de Internet e Correio Eletrônico - Informática para Concursos - Ok ConcursosAline Moura Savassini LucenaNo ratings yet
- Noções de Sistema Operacional - Windows e Linux - Informática para Concursos - Ok ConcursosDocument3 pagesNoções de Sistema Operacional - Windows e Linux - Informática para Concursos - Ok ConcursosAline Moura Savassini LucenaNo ratings yet
- Cir Genital MascDocument56 pagesCir Genital MascWendell Fellipe Baia100% (1)
- 6omcuidar Dos Animais Seg EdDocument20 pages6omcuidar Dos Animais Seg EdAntonio FrescoNo ratings yet
- Regimento Geral Unir 388371390Document68 pagesRegimento Geral Unir 388371390Aline Moura Savassini LucenaNo ratings yet
- Diretrizes Vacinais DiagramadoDocument42 pagesDiretrizes Vacinais DiagramadoAline Moura Savassini LucenaNo ratings yet
- Diretrizes Vacinais - DiagramadoDocument13 pagesDiretrizes Vacinais - DiagramadoLatasha GrahamNo ratings yet
- Diretrizes Vacinais - DiagramadoDocument13 pagesDiretrizes Vacinais - DiagramadoLatasha GrahamNo ratings yet
- Livro Virologia Capc3adtulo 12Document32 pagesLivro Virologia Capc3adtulo 12Aline Moura Savassini LucenaNo ratings yet
- Resumo - LPF 13 - EndocrinoDocument15 pagesResumo - LPF 13 - EndocrinoSarah Soares100% (1)
- Resumo FarmacologiaDocument6 pagesResumo FarmacologiaNICHOLAS VINICIUS SALA DA SILVANo ratings yet
- Climaterio SANARDocument23 pagesClimaterio SANARAna LuizaNo ratings yet
- Monografia TiagoDocument52 pagesMonografia TiagoalisaNo ratings yet
- Apostila 2015Document57 pagesApostila 2015danielNo ratings yet
- Síndrome Do Ovário Policístico - SOPDocument45 pagesSíndrome Do Ovário Policístico - SOPBárbara TauannaNo ratings yet
- Aula 02 HORMÔNIOSDocument17 pagesAula 02 HORMÔNIOSarielly.barbie12No ratings yet
- 2 Fertilidade e Obstetricia em Bovinos - NoakesDocument146 pages2 Fertilidade e Obstetricia em Bovinos - NoakesSílvia CarvalhoNo ratings yet
- Luz para AviculturaDocument38 pagesLuz para AviculturaJoão MariaNo ratings yet
- Resumo Biologia 12ºDocument21 pagesResumo Biologia 12ºMatilde OliveiraNo ratings yet
- Degeneração TesticularDocument14 pagesDegeneração TesticularGabrielle MouraNo ratings yet
- Padrão Hormonal FemininoDocument20 pagesPadrão Hormonal FemininoCENTRO DE TRATAMENTO DA DOR DR. ANDRÉ FRARENo ratings yet
- M1P5Document9 pagesM1P5Daiane Bonini GabbiNo ratings yet
- 3.endocrinologia e Puberdade No MachoDocument55 pages3.endocrinologia e Puberdade No MachoCelly Sousa100% (2)
- 04 - ADOLESCÊNCIA e PUBERDADEDocument6 pages04 - ADOLESCÊNCIA e PUBERDADEAndressa AlbertiNo ratings yet
- Material de Avicultura - Parte 2Document42 pagesMaterial de Avicultura - Parte 2Lhais VitorassiNo ratings yet
- P3F2Document13 pagesP3F2Beatrice MendesNo ratings yet
- Anatomia e Fisiologia Da MulherDocument66 pagesAnatomia e Fisiologia Da MulherRafael Marcos Da SilvaNo ratings yet
- Ciclo MenstrualDocument3 pagesCiclo MenstrualMarina GrisonNo ratings yet
- Fisiologia Da Reproduçao de Bovinos Leiteiros - Aspectos Básicos e ClínicosDocument202 pagesFisiologia Da Reproduçao de Bovinos Leiteiros - Aspectos Básicos e ClínicosDeborahNo ratings yet
- IAMSPE SP 2024 ObjetivamnnDocument45 pagesIAMSPE SP 2024 ObjetivamnnMarco AntonioNo ratings yet
- Endocrinologia ClínicaDocument49 pagesEndocrinologia ClínicaCarolina DonattiNo ratings yet
- GinecologiaDocument549 pagesGinecologiaAntônio Kawakame NetoNo ratings yet
- Reproducao Assistida A3 PDFDocument31 pagesReproducao Assistida A3 PDFMaria LuciaNo ratings yet
- Aula 2. Controle Neuro-Endócrino-Gonadal FêmeaDocument97 pagesAula 2. Controle Neuro-Endócrino-Gonadal Fêmeahelder.siqueiraNo ratings yet
- Apg 29Document13 pagesApg 29Vinicius BarrosNo ratings yet
- Joana Costa, A41284 - Obesidade e InfertilidadeDocument6 pagesJoana Costa, A41284 - Obesidade e Infertilidadea41284No ratings yet
- 23315d-DC Como Avaliar e Tratar Adl Com Puberdade AtrasadaDocument10 pages23315d-DC Como Avaliar e Tratar Adl Com Puberdade AtrasadaAugusto WickertNo ratings yet