You might also like
- Human Chromosomes: An Illustrated Introduction to Human CytogeneticsFrom EverandHuman Chromosomes: An Illustrated Introduction to Human CytogeneticsRating: 5 out of 5 stars5/5 (1)
- Problems of Cell Permeability: International Series of Monographs in Pure and Applied Biology: Modern Trends in Physiological Sciences, Vol. 26From EverandProblems of Cell Permeability: International Series of Monographs in Pure and Applied Biology: Modern Trends in Physiological Sciences, Vol. 26No ratings yet
- DLC PracticalDocument6 pagesDLC PracticalAngoruz Gohain BaruahNo ratings yet
- A Colour Atlas of Haematological Cytology by Hayhoe, PGDocument373 pagesA Colour Atlas of Haematological Cytology by Hayhoe, PGC0ghi86% (7)
- Bookshelf NBK261Document4 pagesBookshelf NBK261nesiaNo ratings yet
- Lab 9Document40 pagesLab 9bob0% (1)
- Lesson 9 Leukocytes1Document2 pagesLesson 9 Leukocytes1Kyla SalgadoNo ratings yet
- Biology HL - Chapter Summaries - Second Edition - Pearson 2014Document34 pagesBiology HL - Chapter Summaries - Second Edition - Pearson 2014jjkjljNo ratings yet
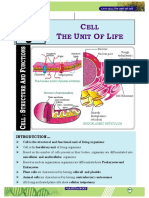
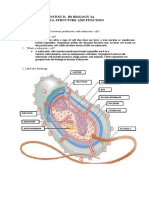
- Cell - The Unit of LifeDocument7 pagesCell - The Unit of LifeAnkan GhoshNo ratings yet
- 1 Biosci The Cells and TissuesDocument7 pages1 Biosci The Cells and TissuesPatricia Rosabelle RoñoNo ratings yet
- Ext - Textbook of Practical Physiology (G.K Pal and Pravati Pal)Document7 pagesExt - Textbook of Practical Physiology (G.K Pal and Pravati Pal)Junaid Shaik100% (1)
- Section - A: Model Paper - 1 Diploma in Medical Lab Technician Second Year Paper - IDocument19 pagesSection - A: Model Paper - 1 Diploma in Medical Lab Technician Second Year Paper - IMohammed MjdNo ratings yet
- Cell Structure and Function WorksheetDocument3 pagesCell Structure and Function WorksheetKiersten RobertsNo ratings yet
- Module (Cell Nucleus)Document12 pagesModule (Cell Nucleus)Edgie JunelaNo ratings yet
- Jovr 04 01 040Document19 pagesJovr 04 01 040JaTi NurwigatiNo ratings yet
- Allot Mindorff Mitosis and MeiosisDocument11 pagesAllot Mindorff Mitosis and MeiosisElTrucoCeroNo ratings yet
- Marieb - CH - 03 - Lecture - Doc (Edited)Document9 pagesMarieb - CH - 03 - Lecture - Doc (Edited)Dustin RamosNo ratings yet
- Fisiología Del Músculo Esquelético: Tema: Introducción Al MionúcleoDocument16 pagesFisiología Del Músculo Esquelético: Tema: Introducción Al MionúcleoDiana Jaramillo HNo ratings yet
- Cell Bio SLDocument103 pagesCell Bio SLkrhimkrNo ratings yet
- Nuclear Pore ComplexesDocument9 pagesNuclear Pore ComplexesMNo ratings yet
- 3 - Normal LeukocytesDocument6 pages3 - Normal LeukocytesMatheus AlmadaNo ratings yet
- Nucleus - Morphology, Structure, Chemical Composition, Functions and Significance of NucleusDocument36 pagesNucleus - Morphology, Structure, Chemical Composition, Functions and Significance of NucleusnegriRDNo ratings yet
- B204 (Cellbio)Document33 pagesB204 (Cellbio)shengsking45No ratings yet
- Blood-Edited 230914 110051Document18 pagesBlood-Edited 230914 110051smpoojasubashNo ratings yet
- Cabulisan, Mildred I.Document8 pagesCabulisan, Mildred I.Mildred Ibe CabulisanNo ratings yet
- Cell DivisionDocument4 pagesCell DivisionAndy AgyemangNo ratings yet
- Nucleus 2015Document24 pagesNucleus 2015Analiz De Jesús LópezNo ratings yet
- Ultrastructure and Epithelial Tissue Histology Prac Notes MEDS2005 2022-2Document89 pagesUltrastructure and Epithelial Tissue Histology Prac Notes MEDS2005 2022-2htparkerrNo ratings yet
- The CellDocument22 pagesThe CellWadabiNo ratings yet
- Part 3 - Laboratory ManualDocument50 pagesPart 3 - Laboratory ManualBianca AnguloNo ratings yet
- Basic Concepts of Bio Chemistry and Cell Biology: Learning ObjectivesDocument52 pagesBasic Concepts of Bio Chemistry and Cell Biology: Learning ObjectivesIyappan SubramaniNo ratings yet
- Activity 3Document3 pagesActivity 3enaniacanoNo ratings yet
- Chapter - 8 Cell As Unit of LifeDocument21 pagesChapter - 8 Cell As Unit of Lifeavishank584No ratings yet
- Cells Review Ws AnswersDocument4 pagesCells Review Ws AnswersTongtun TuntunNo ratings yet
- Pages From 1-Molecular Cell Biology Lodish 5Th EdDocument4 pagesPages From 1-Molecular Cell Biology Lodish 5Th Edapi-3733156100% (1)
- 3 CellsDocument7 pages3 CellsCaviles, Jasmin S.No ratings yet
- Ncert Sol For Cbse Class 9 Sci Chapter 5 The Fundamental Unit of LifeDocument6 pagesNcert Sol For Cbse Class 9 Sci Chapter 5 The Fundamental Unit of LifeShah RukhNo ratings yet
- Exercise-5.1: NCERT Solution For Class 9 Science - Chapter 5 The Fundamental Unit of LifeDocument6 pagesExercise-5.1: NCERT Solution For Class 9 Science - Chapter 5 The Fundamental Unit of Lifenitika chawlaNo ratings yet
- 01 CiliaDocument3 pages01 CiliaLaksh NrusimhanNo ratings yet
- Notes - For - Cell (BOOK)Document10 pagesNotes - For - Cell (BOOK)Macchi MagsNo ratings yet
- Cell The Unit of Life - Old NmodDocument37 pagesCell The Unit of Life - Old NmodShreyaa SiddarthNo ratings yet
- A8 Cell Unit of LifeDocument1 pageA8 Cell Unit of LiferajjoNo ratings yet
- Interphasic NucleusDocument21 pagesInterphasic Nucleushamahamahama3No ratings yet
- Plant Cells Lab ActivityDocument5 pagesPlant Cells Lab ActivityHaris KhanNo ratings yet
- Living Parts: Plant Parts Ofa CellDocument11 pagesLiving Parts: Plant Parts Ofa CellGarvNo ratings yet
- 8 Cell Structure and FunctionDocument21 pages8 Cell Structure and FunctionVince RetutaNo ratings yet
- Brandt 2006Document10 pagesBrandt 2006yalocim666No ratings yet
- Seminar 2 NotesDocument19 pagesSeminar 2 NotesRica Mae A. MarcialNo ratings yet
- APIADOP ANA PHY Lab Activity 2 The Function of Composite CellDocument4 pagesAPIADOP ANA PHY Lab Activity 2 The Function of Composite CellAj ApiadoNo ratings yet
- Cell Structure - Part II Updated 01-11-2016Document47 pagesCell Structure - Part II Updated 01-11-2016Abdulaziz AHNo ratings yet
- Act04 Cell Division - Mitosis DiscussionDocument3 pagesAct04 Cell Division - Mitosis DiscussionPaula Nicole AlanoNo ratings yet
- Plant Animal Cell DiagramsDocument5 pagesPlant Animal Cell Diagramssalagi28No ratings yet
- Bab 1 2 Biomembran PDFDocument84 pagesBab 1 2 Biomembran PDFMuh JufriNo ratings yet
- Biology Chapter 1Document9 pagesBiology Chapter 1Shubham ShawNo ratings yet
- The Cell and Biological Membranes Structure and FunctionDocument9 pagesThe Cell and Biological Membranes Structure and FunctionAvaneesh HNo ratings yet
- BIO Cell Structure Function UnChilled.8532Document9 pagesBIO Cell Structure Function UnChilled.8532Anna WrightNo ratings yet
- CELLS 2.1: Outline: To Give A Brief Account or SummaryDocument10 pagesCELLS 2.1: Outline: To Give A Brief Account or SummaryHrithik SolaniNo ratings yet
- Class Xi Biology Practical Record Writing 2022-23 - Part 3Document54 pagesClass Xi Biology Practical Record Writing 2022-23 - Part 3Om SolankiNo ratings yet
- Research in ProtozoologyFrom EverandResearch in ProtozoologyTze-Tuan ChenRating: 5 out of 5 stars5/5 (1)
- Agilent 385-ELSD Evaporative Light Scattering Detector: For Low Molecular Weight CompoundsDocument8 pagesAgilent 385-ELSD Evaporative Light Scattering Detector: For Low Molecular Weight CompoundsPedro LopesNo ratings yet
- Agilent ELSD - Analysis of Foods ApplicationsDocument12 pagesAgilent ELSD - Analysis of Foods ApplicationsPedro LopesNo ratings yet
- Agilent Openlab CDS - Workstation Requirements and Supported InstrumentsDocument52 pagesAgilent Openlab CDS - Workstation Requirements and Supported InstrumentsPedro LopesNo ratings yet
- Customer ResponsibilitiesDocument5 pagesCustomer ResponsibilitiesPedro LopesNo ratings yet
- User Manual: 1260 Infinity II Isocratic and Quaternary PumpsDocument248 pagesUser Manual: 1260 Infinity II Isocratic and Quaternary PumpsPedro LopesNo ratings yet
- Agilent Best Practices For Using An Agilent LC SystemDocument19 pagesAgilent Best Practices For Using An Agilent LC SystemPedro LopesNo ratings yet
- Agilent 1260 Series (Connect To Thermo MS)Document56 pagesAgilent 1260 Series (Connect To Thermo MS)Pedro LopesNo ratings yet
- Agilent 1200 Infinity - VWD (User Manual)Document244 pagesAgilent 1200 Infinity - VWD (User Manual)Pedro LopesNo ratings yet
- Agilent 1260 Infinity SuppliesDocument2 pagesAgilent 1260 Infinity SuppliesPedro LopesNo ratings yet
- Universidade de Aveiro Departamento de Matem AticaDocument2 pagesUniversidade de Aveiro Departamento de Matem AticaPedro LopesNo ratings yet
- Jurnal Perdata K 1Document3 pagesJurnal Perdata K 1Edi nur HandokoNo ratings yet
- Hankinson - Location Branding - A Study of The Branding Practices of 12 English CitiesDocument16 pagesHankinson - Location Branding - A Study of The Branding Practices of 12 English CitiesNatalia Ney100% (1)
- Aqualab ClinicDocument12 pagesAqualab ClinichonyarnamiqNo ratings yet
- Review Questions & Answers For Midterm1: BA 203 - Financial Accounting Fall 2019-2020Document11 pagesReview Questions & Answers For Midterm1: BA 203 - Financial Accounting Fall 2019-2020Ulaş GüllenoğluNo ratings yet
- Placement TestDocument6 pagesPlacement TestNovia YunitazamiNo ratings yet
- 1-Gaikindo Category Data Jandec2020Document2 pages1-Gaikindo Category Data Jandec2020Tanjung YanugrohoNo ratings yet
- PS4 ListDocument67 pagesPS4 ListAnonymous yNw1VyHNo ratings yet
- 4-Page 7 Ways TM 20Document4 pages4-Page 7 Ways TM 20Jose EstradaNo ratings yet
- Ob AssignmntDocument4 pagesOb AssignmntOwais AliNo ratings yet
- Pilapil v. CADocument2 pagesPilapil v. CAIris Gallardo100% (2)
- PLSQL Day 1Document12 pagesPLSQL Day 1rambabuNo ratings yet
- Post Cold WarDocument70 pagesPost Cold WarZainab WaqarNo ratings yet
- Assignment 2 Malaysian StudiesDocument4 pagesAssignment 2 Malaysian StudiesPenny PunNo ratings yet
- CALIDocument58 pagesCALIleticia figueroaNo ratings yet
- Linkedin Job Invite MessageDocument4 pagesLinkedin Job Invite MessageAxiom IntNo ratings yet
- Feng Shui GeneralDocument36 pagesFeng Shui GeneralPia SalvadorNo ratings yet
- AI in HealthDocument105 pagesAI in HealthxenoachNo ratings yet
- Landslide Hazard Manual: Trainer S HandbookDocument32 pagesLandslide Hazard Manual: Trainer S HandbookMouhammed AbdallahNo ratings yet
- Ikramul (Electrical)Document3 pagesIkramul (Electrical)Ikramu HaqueNo ratings yet
- LR 7833Document11 pagesLR 7833Trung ĐinhNo ratings yet
- ANNAPURNA Sanitary Work 3 FinalDocument34 pagesANNAPURNA Sanitary Work 3 FinalLaxu KhanalNo ratings yet
- 401 Lourdes School Quezon City Inc. v. GarciaDocument2 pages401 Lourdes School Quezon City Inc. v. GarciaIldefonso HernaezNo ratings yet
- 1 Piling LaranganDocument3 pages1 Piling LaranganHannie Jane Salazar HerreraNo ratings yet
- Role of Commodity Exchange in Agricultural GrowthDocument63 pagesRole of Commodity Exchange in Agricultural GrowthSoumyalin Santy50% (2)
- Novedades Jaltest CV en 887Document14 pagesNovedades Jaltest CV en 887Bruce LyndeNo ratings yet
- Concordia: The Lutheran Confessions - ExcerptsDocument39 pagesConcordia: The Lutheran Confessions - ExcerptsConcordia Publishing House28% (25)
- Fort - Fts - The Teacher and ¿Mommy Zarry AdaptaciónDocument90 pagesFort - Fts - The Teacher and ¿Mommy Zarry AdaptaciónEvelin PalenciaNo ratings yet
- Delegated Legislation in India: Submitted ToDocument15 pagesDelegated Legislation in India: Submitted ToRuqaiyaNo ratings yet
- Past Simple of BeDocument2 pagesPast Simple of BeRoxana ClepeNo ratings yet
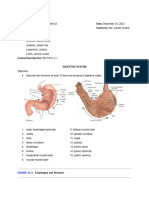
- Digestive System LabsheetDocument4 pagesDigestive System LabsheetKATHLEEN MAE HERMONo ratings yet