You might also like
- Atlas of the Prenatal Mouse BrainFrom EverandAtlas of the Prenatal Mouse BrainUta B. SchambraNo ratings yet
- Cardiogenesis and The Regulation of Cardiac-Specific Gene ExpressionDocument14 pagesCardiogenesis and The Regulation of Cardiac-Specific Gene ExpressiontyasNo ratings yet
- Developmental Dynamics - 2006 - Campione - Cardiovascular Development Toward Biomedical ApplicabilityDocument3 pagesDevelopmental Dynamics - 2006 - Campione - Cardiovascular Development Toward Biomedical Applicabilitystevenburrow06No ratings yet
- Sox2 Acts As A Rheostat of Epithelial To Mesenchymal Transition During Neural Crest DevelopmentDocument12 pagesSox2 Acts As A Rheostat of Epithelial To Mesenchymal Transition During Neural Crest DevelopmentAngelicaNo ratings yet
- Vitalis 2005Document9 pagesVitalis 2005Raj PrabuNo ratings yet
- Ateroesclerosis Kimee NeyssaDocument12 pagesAteroesclerosis Kimee NeyssaNEYSSA KIMEE SAINT VILNo ratings yet
- 56 Koshiba Takeuchi Et AlDocument5 pages56 Koshiba Takeuchi Et AlJamesComeyJustaBitchNo ratings yet
- Reviews: Thyroid Transcription Factors in Development, Differentiation and DiseaseDocument14 pagesReviews: Thyroid Transcription Factors in Development, Differentiation and DiseaseJuan Antonio Herrera LealNo ratings yet
- Historical Perspective: A Decade of Discoveries in Cardiac BiologyDocument8 pagesHistorical Perspective: A Decade of Discoveries in Cardiac BiologyDiego AbrahamNo ratings yet
- The Homeodomain Factor Lbx1 Distinguishes Two Major Programs of Neuronal Differentiation in The Dorsal Spinal CordDocument12 pagesThe Homeodomain Factor Lbx1 Distinguishes Two Major Programs of Neuronal Differentiation in The Dorsal Spinal CordaptureincNo ratings yet
- 1428 FullDocument6 pages1428 FullErik ArturNo ratings yet
- Developmental Biology: The Epicardium in The Embryonic and Adult ZebrafishDocument16 pagesDevelopmental Biology: The Epicardium in The Embryonic and Adult ZebrafishplatusNo ratings yet
- ZhangDocument11 pagesZhangAJMRNo ratings yet
- TMP 8208Document15 pagesTMP 8208FrontiersNo ratings yet
- PAUSED, A Putative Exportin, Acts Pleiotropically in Is Development But Is Dispensable For ViabilityDocument12 pagesPAUSED, A Putative Exportin, Acts Pleiotropically in Is Development But Is Dispensable For ViabilityQuanan HuNo ratings yet
- EmbriologíaDocument16 pagesEmbriologíalucas LópezNo ratings yet
- Publication Congenital Central Hypoventilation Syndrome: Mutation Analysis THE Receptor Tyrosine Kinase RetDocument7 pagesPublication Congenital Central Hypoventilation Syndrome: Mutation Analysis THE Receptor Tyrosine Kinase RetSergioFernandezNo ratings yet
- Mutations in The Very Low-Density Lipoprotein Receptor VLDLR Cause Cerebellar Hypoplasia and Quadrupedal Locomotion in HumansDocument5 pagesMutations in The Very Low-Density Lipoprotein Receptor VLDLR Cause Cerebellar Hypoplasia and Quadrupedal Locomotion in HumanssplichardsNo ratings yet
- Epha4 Preserves Postnatal and Adult Neural Stem Cells in An Undifferentiated State in VivoDocument12 pagesEpha4 Preserves Postnatal and Adult Neural Stem Cells in An Undifferentiated State in VivoFrontiersNo ratings yet
- Lavdas Et Al. - J.neurosc. - 1999 - The MGE Gives Rise To A Population of Early Neurons in The Developing Cerebral CortexDocument8 pagesLavdas Et Al. - J.neurosc. - 1999 - The MGE Gives Rise To A Population of Early Neurons in The Developing Cerebral CortexCris AdFNo ratings yet
- Oct4 and Sox2 - Pluripotency of TheDocument2 pagesOct4 and Sox2 - Pluripotency of TheGelyn Lanuza-AsuncionNo ratings yet
- DescargaDocument18 pagesDescargakirbwer macedoNo ratings yet
- Paper Canty 2009 - PV HolesDocument11 pagesPaper Canty 2009 - PV Holesmaariamunoz9No ratings yet
- Witty Et AlDocument12 pagesWitty Et AlJustin EdwardsNo ratings yet
- 2023 FD CarDocument12 pages2023 FD CarAlejandra Gonzalez DuarteNo ratings yet
- 382 Thyroid Gland Physiology and TestingDocument7 pages382 Thyroid Gland Physiology and TestingJetmoon BlackNo ratings yet
- BDNF Ncbi PDFDocument15 pagesBDNF Ncbi PDFAnand Prakash YadavNo ratings yet
- Heart Malformation Is An Early Response To TCDD in Embryonic ZebrafishDocument10 pagesHeart Malformation Is An Early Response To TCDD in Embryonic ZebrafishIntan Wahyu CahyaniNo ratings yet
- Cooperative Slit and Netrin Signaling Guides Trigeminal Axons Across MidlineDocument19 pagesCooperative Slit and Netrin Signaling Guides Trigeminal Axons Across MidlineMutiara Tri JunisaNo ratings yet
- Genetic Blueprint Cardiac DevelopmentDocument6 pagesGenetic Blueprint Cardiac DevelopmentProf. David Rodriguez MartinezNo ratings yet
- Cripto-1: A Multifunctional Modulator During Embryogenesis and OncogenesisDocument11 pagesCripto-1: A Multifunctional Modulator During Embryogenesis and OncogenesisCristina Romero TrisNo ratings yet
- FT 3351Document10 pagesFT 3351Ningthoujam MeenakshiNo ratings yet
- Baf60mice Takeuchi2007Document7 pagesBaf60mice Takeuchi2007vweakeNo ratings yet
- Cfos, Jun y ExploracionDocument12 pagesCfos, Jun y ExploracionLuisa Fernanda Acevedo ReyesNo ratings yet
- A Spatiotemporal Organ-Wide Gene Expression and Cell Atlas of The Developing Human HeartDocument34 pagesA Spatiotemporal Organ-Wide Gene Expression and Cell Atlas of The Developing Human Heartlei guNo ratings yet
- Familial Neuronal Migration Disorder: Subcortical: Laminar Heterotopia in A Mother and Pachygyria in The SonDocument4 pagesFamilial Neuronal Migration Disorder: Subcortical: Laminar Heterotopia in A Mother and Pachygyria in The SoncthjgfytvfnoNo ratings yet
- Caenorhabditis gonadeDocument21 pagesCaenorhabditis gonadeIstván NagyNo ratings yet
- The Drosophila Melanogaster Circadian Pacemaker Circuit: Review ArticleDocument9 pagesThe Drosophila Melanogaster Circadian Pacemaker Circuit: Review ArticleAli TarkashvandNo ratings yet
- Development of The Vertebrate Heart: Mabelle Ashe and David Bader, PHDDocument2 pagesDevelopment of The Vertebrate Heart: Mabelle Ashe and David Bader, PHDTariq MuneerNo ratings yet
- Developmental Dynamics - 2000 - Lough - Endoderm and Heart DevelopmentDocument16 pagesDevelopmental Dynamics - 2000 - Lough - Endoderm and Heart DevelopmentMayu PatNo ratings yet
- Proche L Sanchez Zoo L 03Document12 pagesProche L Sanchez Zoo L 03rustyjuuNo ratings yet
- YangDocument13 pagesYangAJMRNo ratings yet
- Lambert & Goffinet - Biochem - Pharmac. - 1998 - A New View of Early Corticla DevelopmentDocument7 pagesLambert & Goffinet - Biochem - Pharmac. - 1998 - A New View of Early Corticla DevelopmentCris AdFNo ratings yet
- DNA methylation regulates heart development genesDocument21 pagesDNA methylation regulates heart development genesAJNo ratings yet
- 10 FullDocument9 pages10 FullDokdem AjaNo ratings yet
- Top 4 Genetics PapersDocument139 pagesTop 4 Genetics PapersManjunath.RNo ratings yet
- 1097 0185 (20000701) 259 3 248 Aid Ar30 3.0Document15 pages1097 0185 (20000701) 259 3 248 Aid Ar30 3.0Odrig De LavalleNo ratings yet
- 6142 FullDocument12 pages6142 FullAmol MundeNo ratings yet
- Shapiro 1997Document6 pagesShapiro 1997JohnnyNo ratings yet
- Pattern Formation and Growth During Floral OrganogDocument9 pagesPattern Formation and Growth During Floral OrganogSweeti MandalNo ratings yet
- 4462 FullDocument10 pages4462 FullDokdem AjaNo ratings yet
- Developmental expression of tshz2 and tshz3b in zebrafishDocument9 pagesDevelopmental expression of tshz2 and tshz3b in zebrafishFernando CasaresNo ratings yet
- Schridde2006 IhDocument13 pagesSchridde2006 Ihiulia andreeaNo ratings yet
- A Review of Disrupted-in-Schizophrenia-1 (Disc1) : Neurodevelopment, Cognition, and Mental ConditionsDocument9 pagesA Review of Disrupted-in-Schizophrenia-1 (Disc1) : Neurodevelopment, Cognition, and Mental Conditionsrocambolescas perthNo ratings yet
- LIF Transduces Contradictory Signals On Capillary Outgrowth Through Induction of Stat3 and MAP KinaseDocument9 pagesLIF Transduces Contradictory Signals On Capillary Outgrowth Through Induction of Stat3 and MAP Kinasetele_noteNo ratings yet
- Genotype-Phenotype Correlation, and Phenotypic HeterogeneityDocument9 pagesGenotype-Phenotype Correlation, and Phenotypic Heterogeneitykorokoro12No ratings yet
- Cells of The Adult Human Heart: ArticleDocument34 pagesCells of The Adult Human Heart: ArticleJoão VítorNo ratings yet
- Structure and Function of The Developing Zebrafish HeartDocument10 pagesStructure and Function of The Developing Zebrafish HeartKarthik ShettyNo ratings yet
- 5865 FullDocument14 pages5865 Fullmsaponaro6ࢮNo ratings yet
- Norepinephrine-Deficient Mice Have Increased Susceptibility To Seizure-Inducing StimuliDocument8 pagesNorepinephrine-Deficient Mice Have Increased Susceptibility To Seizure-Inducing StimuliFrontiersNo ratings yet
- Healty RiskDocument7 pagesHealty RiskkianaNo ratings yet
- Groups Count Sum AverageDocument2 pagesGroups Count Sum AveragekianaNo ratings yet
- Suhu tubuh dan regulasi termalDocument7 pagesSuhu tubuh dan regulasi termalNurul Asri MulyaniNo ratings yet
- Gata6 PDFDocument9 pagesGata6 PDFkianaNo ratings yet
- STR BaseDocument3 pagesSTR BasekianaNo ratings yet
- HAL Cycadopsida Ginkgopsida Pinopsida Gnetopsida: Habitus DaunDocument2 pagesHAL Cycadopsida Ginkgopsida Pinopsida Gnetopsida: Habitus DaunkianaNo ratings yet
- Ventrikel FormationDocument14 pagesVentrikel FormationkianaNo ratings yet
- Gambar Klasifikasi Kelinci Lop: KingdomDocument4 pagesGambar Klasifikasi Kelinci Lop: Kingdomkiana100% (1)
- Silverthorn ImunDocument3 pagesSilverthorn ImunkianaNo ratings yet
- Radiology, Anesthesia, Gynecology NeetDocument71 pagesRadiology, Anesthesia, Gynecology NeetDivya Joyce100% (1)
- NPE Teaching Manual El-Khuffash 2019 - Dr. Ahmad Kautsar, Sp.ADocument117 pagesNPE Teaching Manual El-Khuffash 2019 - Dr. Ahmad Kautsar, Sp.AlaurentiaNo ratings yet
- Cape Biology Unit 2 Module 2 MCQDocument16 pagesCape Biology Unit 2 Module 2 MCQwilliam bridgemohansinghNo ratings yet
- Ec1001 - Medical Electronics - Question Bank (2marks With Answer)Document26 pagesEc1001 - Medical Electronics - Question Bank (2marks With Answer)deepanece100% (1)
- Acute Biological Crisis: Louis Carlos O. RoderosDocument76 pagesAcute Biological Crisis: Louis Carlos O. RoderosLouis RoderosNo ratings yet
- Ebstein Anomaly:: Fits Best With This Clinical Picture?Document5 pagesEbstein Anomaly:: Fits Best With This Clinical Picture?Galaleldin AliNo ratings yet
- 1BI0 2H Mark-Scheme 20180822Document25 pages1BI0 2H Mark-Scheme 20180822Xiao ShadowlordNo ratings yet
- Shock An OverviewDocument41 pagesShock An OverviewelaineNo ratings yet
- The Heart Anatomy and PhysiologyDocument15 pagesThe Heart Anatomy and PhysiologyAmelie Sta RomanaNo ratings yet
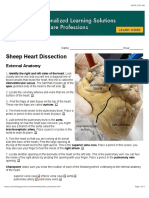
- Sheep Heart Dissection GuideDocument3 pagesSheep Heart Dissection GuideMorgan DanielsNo ratings yet
- GD 10 Biology Life Processes Transportation Ref NotesDocument5 pagesGD 10 Biology Life Processes Transportation Ref Notesaloma.alvaresNo ratings yet
- AIPMT 2013 Code W1 Question PaperDocument43 pagesAIPMT 2013 Code W1 Question PaperNageswarNo ratings yet
- Pediatric and Neonatal Respiratory Care DevelopmentDocument205 pagesPediatric and Neonatal Respiratory Care DevelopmentAlexander Santiago ParelNo ratings yet
- Grade 7 CH 4.05 The HeartDocument12 pagesGrade 7 CH 4.05 The HeartMunmun AfnanNo ratings yet
- B4 - Organising Animals and PlantsDocument4 pagesB4 - Organising Animals and PlantsozmanNo ratings yet
- Fluid Status and Fluid Responsiveness PDFDocument8 pagesFluid Status and Fluid Responsiveness PDFGiselle BaiãoNo ratings yet
- MS Wrep IIDocument51 pagesMS Wrep IIiana-almocera-6970No ratings yet
- Circulatory System Lesson Explains How Heart and Blood Vessels WorkDocument48 pagesCirculatory System Lesson Explains How Heart and Blood Vessels WorkNaisy Magalona100% (2)
- Heart & Hemodynamics NotesDocument8 pagesHeart & Hemodynamics NotesBrandice BradleyNo ratings yet
- Compare and Contrast Process in Plants and AnimalsDocument8 pagesCompare and Contrast Process in Plants and AnimalsKimberly EamilaoNo ratings yet
- IJSO Question Paper 2007 (Theory)Document14 pagesIJSO Question Paper 2007 (Theory)Science Olympiad BlogNo ratings yet
- Cardiodynamics - Dra-1. ValerioDocument7 pagesCardiodynamics - Dra-1. ValerioJorelyn FriasNo ratings yet
- Lecture On Cardiac Cycle by DR RoomiDocument43 pagesLecture On Cardiac Cycle by DR RoomiMudassar Roomi100% (2)
- How To Dissect RatDocument30 pagesHow To Dissect RatHend MaarofNo ratings yet
- Conginital Heart DiseaseDocument19 pagesConginital Heart DiseaseSanthosh.S.UNo ratings yet
- Multiple Choice Questions (MCQ) Topic Quiz Cardiovascular SystemDocument19 pagesMultiple Choice Questions (MCQ) Topic Quiz Cardiovascular SystemMarimuthu ANo ratings yet
- Cardiovascular Disease in Small Animal Medicine (VetBooks - Ir) PDFDocument397 pagesCardiovascular Disease in Small Animal Medicine (VetBooks - Ir) PDFGrisel Leal GomezNo ratings yet
- Bainbridge ReflexDocument4 pagesBainbridge ReflexJoshua EspinosaNo ratings yet
- The Heart: Structure and Function in 40 CharactersDocument3 pagesThe Heart: Structure and Function in 40 CharactersTharanga HewabuhageNo ratings yet
- Ob Peds Lecture Notes Nurse NursingDocument70 pagesOb Peds Lecture Notes Nurse Nursinglilchibaby3161100% (2)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 3.5 out of 5 stars3.5/5 (2)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessFrom Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessRating: 4 out of 5 stars4/5 (33)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceFrom EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceRating: 4.5 out of 5 stars4.5/5 (515)
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindFrom EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindRating: 4.5 out of 5 stars4.5/5 (93)
- Crypt: Life, Death and Disease in the Middle Ages and BeyondFrom EverandCrypt: Life, Death and Disease in the Middle Ages and BeyondRating: 4 out of 5 stars4/5 (3)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- This Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyFrom EverandThis Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyRating: 3.5 out of 5 stars3.5/5 (31)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionFrom EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionRating: 4 out of 5 stars4/5 (811)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesFrom EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesRating: 4.5 out of 5 stars4.5/5 (397)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorFrom EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNo ratings yet
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldFrom EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldRating: 4.5 out of 5 stars4.5/5 (18)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedFrom EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedRating: 4 out of 5 stars4/5 (11)
- The Lives of Bees: The Untold Story of the Honey Bee in the WildFrom EverandThe Lives of Bees: The Untold Story of the Honey Bee in the WildRating: 4.5 out of 5 stars4.5/5 (44)
- The Second Brain: A Groundbreaking New Understanding of Nervous Disorders of the Stomach and IntestineFrom EverandThe Second Brain: A Groundbreaking New Understanding of Nervous Disorders of the Stomach and IntestineRating: 4 out of 5 stars4/5 (17)
- Eels: An Exploration, from New Zealand to the Sargasso, of the World's Most Mysterious FishFrom EverandEels: An Exploration, from New Zealand to the Sargasso, of the World's Most Mysterious FishRating: 4 out of 5 stars4/5 (30)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsFrom EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsRating: 4.5 out of 5 stars4.5/5 (4)
- Human Errors: A Panorama of Our Glitches, from Pointless Bones to Broken GenesFrom EverandHuman Errors: A Panorama of Our Glitches, from Pointless Bones to Broken GenesRating: 3.5 out of 5 stars3.5/5 (56)
- The Mind & The Brain: Neuroplasticity and the Power of Mental ForceFrom EverandThe Mind & The Brain: Neuroplasticity and the Power of Mental ForceNo ratings yet
- Gathering Moss: A Natural and Cultural History of MossesFrom EverandGathering Moss: A Natural and Cultural History of MossesRating: 4.5 out of 5 stars4.5/5 (347)
- The Dog Who Couldn't Stop Loving: How Dogs Have Captured Our Hearts for Thousands of YearsFrom EverandThe Dog Who Couldn't Stop Loving: How Dogs Have Captured Our Hearts for Thousands of YearsNo ratings yet
- Younger for Life: Feel Great and Look Your Best with the New Science of AutojuvenationFrom EverandYounger for Life: Feel Great and Look Your Best with the New Science of AutojuvenationRating: 4 out of 5 stars4/5 (1)
- Darwin's Dangerous Idea: Evolution and the Meaning of LifeFrom EverandDarwin's Dangerous Idea: Evolution and the Meaning of LifeRating: 4 out of 5 stars4/5 (523)
- Superlative: The Biology of ExtremesFrom EverandSuperlative: The Biology of ExtremesRating: 4.5 out of 5 stars4.5/5 (51)
- Why We Sleep: Unlocking the Power of Sleep and DreamsFrom EverandWhy We Sleep: Unlocking the Power of Sleep and DreamsRating: 4.5 out of 5 stars4.5/5 (2083)
- Lymph & Longevity: The Untapped Secret to HealthFrom EverandLymph & Longevity: The Untapped Secret to HealthRating: 4.5 out of 5 stars4.5/5 (13)