You might also like
- Effect of ammonia concentration on rumen microbial protein productionDocument10 pagesEffect of ammonia concentration on rumen microbial protein productionHidayat Anjar AsmaraniNo ratings yet
- Lo Del Almidon 1990Document5 pagesLo Del Almidon 1990Alessandra CalderonNo ratings yet
- Estudos de Saccharomyces Cerevisiae Imobilizados. Análise de Fermentação Rápida Contínua de Etanol em Reator de Células ImobilizadasDocument15 pagesEstudos de Saccharomyces Cerevisiae Imobilizados. Análise de Fermentação Rápida Contínua de Etanol em Reator de Células ImobilizadasMarina Cunha SiebenNo ratings yet
- Fermentation 04 00079Document9 pagesFermentation 04 00079CHIRANJEEVINo ratings yet
- Hydrolisis and FlavorDocument13 pagesHydrolisis and FlavorSara PalermoNo ratings yet
- Measuring milk's reducing capacity through iodometric titrationDocument8 pagesMeasuring milk's reducing capacity through iodometric titrationAFDHAL PASANo ratings yet
- Importance of Specific Growth Rate For Subtilisin Expression in Fed-Batch Cultivation of Bacillus Subtilis Spoiig MutantDocument5 pagesImportance of Specific Growth Rate For Subtilisin Expression in Fed-Batch Cultivation of Bacillus Subtilis Spoiig MutantDavid SantiagoNo ratings yet
- A Note On Quantitative Urobilinogen Determinations: Bernard BalikovDocument5 pagesA Note On Quantitative Urobilinogen Determinations: Bernard BalikovTanveerNo ratings yet
- Biotech Bioengineering - 20 April 1987 - Bar - An Unusual Pattern of Product Inhibition Batch Acetic Acid FermentationDocument3 pagesBiotech Bioengineering - 20 April 1987 - Bar - An Unusual Pattern of Product Inhibition Batch Acetic Acid FermentationEvelin RamirezNo ratings yet
- Aiba 1968 Kinetics of Product Inhibition in Alcohol FermentationDocument20 pagesAiba 1968 Kinetics of Product Inhibition in Alcohol Fermentationrishu2525No ratings yet
- B Subtilis Fed Batch AmylaseDocument7 pagesB Subtilis Fed Batch AmylasemrkanaNo ratings yet
- Enhanced Cellulase Production in Fed-Batch Solid State Fermentation of SL-1Document4 pagesEnhanced Cellulase Production in Fed-Batch Solid State Fermentation of SL-1Mella Rosa SebiferaNo ratings yet
- Protein Isolation TechniquesDocument8 pagesProtein Isolation TechniquesIsla CalugayNo ratings yet
- Influence of PH and Ionic Environment On Thermal Aggregation of Whey ProteinsDocument5 pagesInfluence of PH and Ionic Environment On Thermal Aggregation of Whey ProteinsJohnatan HubsNo ratings yet
- Whey Protein Related FactorsDocument6 pagesWhey Protein Related FactorsNGUYỄN THỊ THANH MINH阮氏青明No ratings yet
- Protease Production From Bacillus SubtilisDocument27 pagesProtease Production From Bacillus Subtilisanon_biochemistrystudentNo ratings yet
- Alcoholic Fermentation With Different Initial Glucose Concentration Using Immobilized Yeast in Calcium Alginate GelDocument7 pagesAlcoholic Fermentation With Different Initial Glucose Concentration Using Immobilized Yeast in Calcium Alginate GelViệt Euro ĐinhNo ratings yet
- Fermentation Rates of Maltose and Glucose in Alkaline SolutionsDocument7 pagesFermentation Rates of Maltose and Glucose in Alkaline Solutionsgibrambo5770No ratings yet
- QUESTION Bank Biochemical EngineeringDocument4 pagesQUESTION Bank Biochemical EngineeringMilito GilbertoNo ratings yet
- Ethanol Production From WheyDocument4 pagesEthanol Production From WheySalman Zafar100% (1)
- Tea Leaf Polyphenol OxidaseDocument10 pagesTea Leaf Polyphenol OxidaseHồng TrinhNo ratings yet
- Official: 2021 Microbial Enumeration Tests-Nutritional and Dietary SupplementsDocument6 pagesOfficial: 2021 Microbial Enumeration Tests-Nutritional and Dietary SupplementsselviniaNo ratings yet
- Mechanism of Cytoplasmatic PH Regulation in Hypoxic Maize Root Tips and Its Role in Survival Under HypoxiaDocument5 pagesMechanism of Cytoplasmatic PH Regulation in Hypoxic Maize Root Tips and Its Role in Survival Under HypoxiaGabriel ZahariaNo ratings yet
- Estimating Proteins Using a Micro-Biuret MethodDocument13 pagesEstimating Proteins Using a Micro-Biuret MethodVõ Huy HoàngNo ratings yet
- Exercise 5.1 & 5.2 Techniques For Protein Analysis Protein Isolation & Determination of Protein Concentration by SpectrophotometryDocument7 pagesExercise 5.1 & 5.2 Techniques For Protein Analysis Protein Isolation & Determination of Protein Concentration by SpectrophotometryJulie Ann Estaras FelicesNo ratings yet
- Efficiency of Fatty Acid Synthesis byDocument6 pagesEfficiency of Fatty Acid Synthesis byKatherine GarciaNo ratings yet
- Pichia FermentationDocument11 pagesPichia FermentationmicromanpNo ratings yet
- Effect of Yeast Extract On Alpha-Amylase Synthesis by Bacillus AmyloliquefaciensDocument6 pagesEffect of Yeast Extract On Alpha-Amylase Synthesis by Bacillus AmyloliquefaciensAndalib TabassumNo ratings yet
- Laboratory Report Pharmaceutical Biotechnology: National University of Ho Chi Minh City International UniversityDocument19 pagesLaboratory Report Pharmaceutical Biotechnology: National University of Ho Chi Minh City International UniversityAn Ngoc HoaiNo ratings yet
- Analysis of Wheat Germ Acid Phosphatase Lab Week 6Document5 pagesAnalysis of Wheat Germ Acid Phosphatase Lab Week 6Jay PatelNo ratings yet
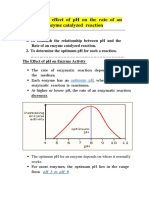
- The Effect of pH on Enzyme ActivityDocument12 pagesThe Effect of pH on Enzyme ActivityAb AbNo ratings yet
- Effect of Modified Serum Immunoglobulins: Kaolin TreatmentDocument4 pagesEffect of Modified Serum Immunoglobulins: Kaolin TreatmentAmit KumarNo ratings yet
- Buffered Peptone Water (Iso) PH 7.0 (9262) : Revision: 6 Effective Date: 4/5/2017Document3 pagesBuffered Peptone Water (Iso) PH 7.0 (9262) : Revision: 6 Effective Date: 4/5/2017nurullahfatihNo ratings yet
- Continuous Fermentation To Produce Fungal Yield and Chemical Composition of Protein. Effect Growth Rate On The BiomassDocument9 pagesContinuous Fermentation To Produce Fungal Yield and Chemical Composition of Protein. Effect Growth Rate On The Biomassandrea galibay perezNo ratings yet
- Urease Activity in Soybean MealDocument4 pagesUrease Activity in Soybean MealAJ QuintoNo ratings yet
- Cinetica Fermentacion 1Document21 pagesCinetica Fermentacion 1Santiago OrtizNo ratings yet
- 1999) Optimization of L-Lactic Acid Feeding For The Production of Poly-D-3-Hydroxybutyric Acid by Alcaligenes Eutrophus in Fed-BatchDocument6 pages1999) Optimization of L-Lactic Acid Feeding For The Production of Poly-D-3-Hydroxybutyric Acid by Alcaligenes Eutrophus in Fed-Batchadmos0319No ratings yet
- Laboratory Analytical Procedure: Chemical Analysis and Testing TaskDocument8 pagesLaboratory Analytical Procedure: Chemical Analysis and Testing TaskJuan Alzate JaramilloNo ratings yet
- Isolation of Invertase PHDocument4 pagesIsolation of Invertase PHJohn Mark Flores VillenaNo ratings yet
- FACTORS AFFECTING THE ASSAY OF HYALURONIDASEDocument15 pagesFACTORS AFFECTING THE ASSAY OF HYALURONIDASEDat VuNo ratings yet
- Aem00187 0216Document7 pagesAem00187 0216MARIA DE LA PAZ GRAJEDA PINEDANo ratings yet
- Características Del Aceite Extraído Con Fosfatasa Recombinante LIS KATERINEDocument6 pagesCaracterísticas Del Aceite Extraído Con Fosfatasa Recombinante LIS KATERINEYury MartinezNo ratings yet
- Physico-Chemical Properties of MilkDocument45 pagesPhysico-Chemical Properties of MilkMohammad Ashraf Paul100% (2)
- Angiotensin I-Converting Enzyme Inhibitory Peptides Isolated From Tofuyo Fermented Soybean FoodDocument6 pagesAngiotensin I-Converting Enzyme Inhibitory Peptides Isolated From Tofuyo Fermented Soybean FoodPablo GabrielNo ratings yet
- LAB_DEMO_LOWRYS_METHODDocument20 pagesLAB_DEMO_LOWRYS_METHODM. GokulakannanNo ratings yet
- Enzymatic Modification of Egg Yolk EmulsionsDocument8 pagesEnzymatic Modification of Egg Yolk EmulsionsLuis LamadridNo ratings yet
- Manufacture Yogurt. Optimal Conditions and Kinetics The Prefermentation ProcessDocument19 pagesManufacture Yogurt. Optimal Conditions and Kinetics The Prefermentation ProcessNorma JenarezNo ratings yet
- Sodium Lactate Antimicrobial ActiveDocument8 pagesSodium Lactate Antimicrobial ActiveEndang Asih Safitri100% (1)
- Fungal EnzymesDocument9 pagesFungal EnzymesYui Bioscience PalsanNo ratings yet
- Continuous Fermentation of Sago Starch to Solvent Using C. saccharobutylicumDocument9 pagesContinuous Fermentation of Sago Starch to Solvent Using C. saccharobutylicumputrianneNo ratings yet
- 1 s2.0 S002203021100213X MainDocument9 pages1 s2.0 S002203021100213X MainJuniClaudia13No ratings yet
- Proteas ADocument20 pagesProteas AIvone Yanira ChoqueticoNo ratings yet
- I. The Role of Pyruvate As An Intermediate Bruno Rosenfeld Ernst SimonDocument11 pagesI. The Role of Pyruvate As An Intermediate Bruno Rosenfeld Ernst SimonreynaldiNo ratings yet
- PhD Qualifier Questions: Bioprocess EngineeringDocument3 pagesPhD Qualifier Questions: Bioprocess EngineeringJustin DeVitoNo ratings yet
- 0065 2571 (72) 90019 2Document20 pages0065 2571 (72) 90019 2Muhammad Akbar SusenoNo ratings yet
- Lsochrysis Galbana: Optimal Growth Conditions ForDocument12 pagesLsochrysis Galbana: Optimal Growth Conditions ForHuy Hữu NguyễnNo ratings yet
- Co- and Post-Translational Modifications of Therapeutic Antibodies and ProteinsFrom EverandCo- and Post-Translational Modifications of Therapeutic Antibodies and ProteinsNo ratings yet
- Plant and Animal Bio-Chemistry - Including Information on Amino Acids, Proteins, Pigments and Other Chemical Constituents of Organic MatterFrom EverandPlant and Animal Bio-Chemistry - Including Information on Amino Acids, Proteins, Pigments and Other Chemical Constituents of Organic MatterNo ratings yet
- As NZS 1865 1997 Aluminium and Aluminium Alloys Drawn Wire Rod Bar and StripDocument7 pagesAs NZS 1865 1997 Aluminium and Aluminium Alloys Drawn Wire Rod Bar and StripYasser Hammad MohamedNo ratings yet
- Triple Science Chemistry Chemical Analysis Revision QuestionsDocument32 pagesTriple Science Chemistry Chemical Analysis Revision QuestionsShakira MaribaoNo ratings yet
- P K Nag SolutionDocument5 pagesP K Nag SolutionMuthu Ezhilan80% (5)
- Assignment 5: Unit 7 - Week 5Document1 pageAssignment 5: Unit 7 - Week 5atulNo ratings yet
- General PHARMAKOLOGY - Загальна фармакологія - Курс лекцій - изд.2005-215 сDocument219 pagesGeneral PHARMAKOLOGY - Загальна фармакологія - Курс лекцій - изд.2005-215 сSahil Saini100% (4)
- Design & Analysis of A Production Well in An Offshore Production FeildDocument26 pagesDesign & Analysis of A Production Well in An Offshore Production FeildAnonymous 1MjHWwNo ratings yet
- Nama: Mochamad Rifli Al Rahman NIM: 24030118130092 Tugas: Kimia Anorganik IvDocument6 pagesNama: Mochamad Rifli Al Rahman NIM: 24030118130092 Tugas: Kimia Anorganik IvMochamad Rifli Al RahmanNo ratings yet
- TiO2 (RC 635)Document1 pageTiO2 (RC 635)medo.k016No ratings yet
- Genapol PF 10Document2 pagesGenapol PF 10Mohamed HalemNo ratings yet
- Medicina Mitocondrial Vol 1Document485 pagesMedicina Mitocondrial Vol 1ArianPedrozaNo ratings yet
- Chemistry Final Cheat SheetDocument1 pageChemistry Final Cheat SheetScott Allred100% (2)
- HS-800 2C Technical Data SheetDocument3 pagesHS-800 2C Technical Data SheetĐạt LêNo ratings yet
- Earth's AtmosphereDocument7 pagesEarth's AtmosphereKristine CastleNo ratings yet
- MKS-112-TS-AZ7-RPT-03 - A - Design Report For Internal Corrosion Monitoring SystemDocument16 pagesMKS-112-TS-AZ7-RPT-03 - A - Design Report For Internal Corrosion Monitoring SystemĐiệnBiênNhâm100% (1)
- 25X5 GI Strip - Data SheetDocument1 page25X5 GI Strip - Data SheetSandeep VijayakumarNo ratings yet
- Heat Transfer and Variable Loads Sept 2017 Rev 6 PresentationDocument49 pagesHeat Transfer and Variable Loads Sept 2017 Rev 6 PresentationFritzel OyaoNo ratings yet
- Audit Criteria: AC7114/1S REV. MDocument47 pagesAudit Criteria: AC7114/1S REV. MCaio Fazzioli TavaresNo ratings yet
- Niosh SHO Exam Notes - Module 3 Question and Answer PDFDocument19 pagesNiosh SHO Exam Notes - Module 3 Question and Answer PDFExcellentdeals4all94% (51)
- 9 - P-Block Elements PDFDocument27 pages9 - P-Block Elements PDFthinkiit86% (7)
- Metallurgy For Non Metallurgist - Module-2Document65 pagesMetallurgy For Non Metallurgist - Module-2ajaysharma_1009No ratings yet
- Porphyrins PDFDocument17 pagesPorphyrins PDFPk JaatNo ratings yet
- ANSWER MPSM Johor Kimia Kertas 3 Set 2 2018 PDFDocument8 pagesANSWER MPSM Johor Kimia Kertas 3 Set 2 2018 PDFSiti Hajar Abd HamidNo ratings yet
- Pressure Swing Adsorption in The Unit Operations LaboratoryDocument8 pagesPressure Swing Adsorption in The Unit Operations Laboratorydilip matalNo ratings yet
- Determining salt content in dough and breadDocument2 pagesDetermining salt content in dough and breadNiels YOUSSANo ratings yet
- Iriotec - 8850 - Merck - TDS (For Rest of The World) PDFDocument2 pagesIriotec - 8850 - Merck - TDS (For Rest of The World) PDFxy2zjgNo ratings yet
- Material SpecificationDocument5 pagesMaterial SpecificationMuthu GaneshNo ratings yet
- Definition of TermsDocument6 pagesDefinition of TermsFrancisco ZieNo ratings yet
- USEPA, Wet Scrubber Inspection and Evaluation Manual (Sep. 1983) PDFDocument164 pagesUSEPA, Wet Scrubber Inspection and Evaluation Manual (Sep. 1983) PDFChristopher Brown100% (1)
- Summary of Scigress Commands Practical 1: Task Scigress Command Shortcut Comments/notesDocument4 pagesSummary of Scigress Commands Practical 1: Task Scigress Command Shortcut Comments/notesnoorNo ratings yet
- Kloppers Critical 2003 PDFDocument363 pagesKloppers Critical 2003 PDF조기현No ratings yet