You might also like
- Unidade Motora e Suas SindromesDocument39 pagesUnidade Motora e Suas SindromesCibele BragaNo ratings yet
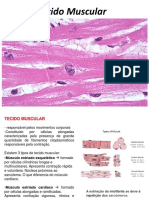
- Tecido MuscularDocument2 pagesTecido MuscularJULIANA GONÇALVES SAMPAIONo ratings yet
- Tecido MuscularDocument5 pagesTecido MuscularRodrigo SousaNo ratings yet
- Tecido Muscular - ResumoDocument6 pagesTecido Muscular - ResumoHelena MarcondesNo ratings yet
- Estudo Dirigido 2Document19 pagesEstudo Dirigido 2moematimoNo ratings yet
- HistologiaPL - Tecido MuscularDocument35 pagesHistologiaPL - Tecido Muscularviolleta gomia100% (1)
- Fisiologia MuscularDocument17 pagesFisiologia MuscularMaria Baião BasílioNo ratings yet
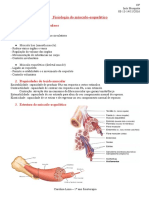
- Sistema Muscular - FisiologiaDocument5 pagesSistema Muscular - FisiologiaKauan GabrielNo ratings yet
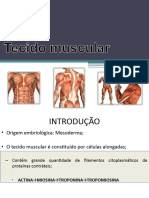
- Tecido MuscularDocument4 pagesTecido MuscularDeadmanNo ratings yet
- Tecido MuscularDocument9 pagesTecido MuscularIsadora RuthNo ratings yet
- OpenAccess Lorente 9788580393996 08Document12 pagesOpenAccess Lorente 9788580393996 08Larissa Medicina veterináriaNo ratings yet
- Sistema Músculo-EsqueléticoDocument21 pagesSistema Músculo-EsqueléticoiotticarolNo ratings yet
- Resumo Tecido MuscularDocument5 pagesResumo Tecido MuscularLaryssa AmorimNo ratings yet
- Fisiologia Da Contração Muscular - AulaDocument6 pagesFisiologia Da Contração Muscular - AulaISADHORA MARIA MARAN DE SOUZANo ratings yet
- Tecido MuscularDocument85 pagesTecido MuscularLarissa MendesNo ratings yet
- Medresumos 2016 - OmfDocument16 pagesMedresumos 2016 - OmfMirianNo ratings yet
- Tecido MuscularDocument5 pagesTecido MusculargabrieltavaresNo ratings yet
- Tecido Muscular - RESUMODocument3 pagesTecido Muscular - RESUMOJuliano JoaquimNo ratings yet
- Tecido MuscularDocument6 pagesTecido Muscularlau24bronzattoNo ratings yet
- Músculos e Receptores MuscularesDocument6 pagesMúsculos e Receptores MuscularesLuciani MoreiraNo ratings yet
- #Resumo#Document10 pages#Resumo#Monitoriadefisio100% (2)
- Resumo Tecido Muscular - JunqueiraDocument4 pagesResumo Tecido Muscular - JunqueiraLucas Thiago BorgesNo ratings yet
- FisiologiaDocument120 pagesFisiologiaMagno DuarteNo ratings yet
- 13 Tec MuscularDocument47 pages13 Tec MuscularmalubastosNo ratings yet
- Contração Muscular - SarcômeroDocument7 pagesContração Muscular - SarcômeroJoyceMedeirosNo ratings yet
- Fisiologia Da Contrac3a7c3a3o Muscular - RoDocument36 pagesFisiologia Da Contrac3a7c3a3o Muscular - RoMarcelo CostaNo ratings yet
- Sistema MuscularDocument23 pagesSistema MuscularWarleyNo ratings yet
- Histologia Do Tecido Muscular @GusthaDiasDocument3 pagesHistologia Do Tecido Muscular @GusthaDiasAna Clara FrotaNo ratings yet
- Bioquímica Da Força e Contração MuscularDocument10 pagesBioquímica Da Força e Contração MuscularhellensabinoNo ratings yet
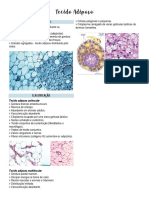
- Tecido MuscularDocument3 pagesTecido MuscularRaquel ChequeleiroNo ratings yet
- Tecido MuscularDocument4 pagesTecido MuscularMirelleNo ratings yet
- Resumo Sanar - Fisiologia Sistema Muscular ÓsseoDocument37 pagesResumo Sanar - Fisiologia Sistema Muscular ÓsseoestudiosojulianoNo ratings yet
- Fisiologia Do Musc EsqueléticoDocument32 pagesFisiologia Do Musc EsqueléticosalvadoorjrNo ratings yet
- Tecido MuscularDocument6 pagesTecido MuscularmayraNo ratings yet
- Junção NeuroDocument33 pagesJunção NeuroFafnir Kesk100% (2)
- Aula 021659031827Document12 pagesAula 021659031827Jurandi AraujoNo ratings yet
- Apg 20 - Semestre 2Document4 pagesApg 20 - Semestre 2Bruna BuenoNo ratings yet
- Cabulas Dos Musculos 1Document1 pageCabulas Dos Musculos 1api-26429188No ratings yet
- Histologia Tecido MuscularDocument5 pagesHistologia Tecido Muscularmariany100% (2)
- Fisiologia Do Músculo-EsqueléticoDocument12 pagesFisiologia Do Músculo-EsqueléticoAna CostaNo ratings yet
- Sistema MuscularDocument18 pagesSistema MuscularGessica Fabiana Cardoso0% (1)
- Contração MuscularDocument7 pagesContração MuscularBeatriz Teixeira MonteiroNo ratings yet
- Tecido Muscular PDFDocument2 pagesTecido Muscular PDFvinimarciaNo ratings yet
- Fechamento Tutoria 12Document12 pagesFechamento Tutoria 12vanessa da silva caetanoNo ratings yet
- Fisiologia Do Músculo-EsqueléticoDocument12 pagesFisiologia Do Músculo-EsqueléticoAna CostaNo ratings yet
- M10P2 - Sistema Motor SomáticoDocument8 pagesM10P2 - Sistema Motor Somáticoana flavia furtado telesNo ratings yet
- Tecido MuscularDocument7 pagesTecido MuscularRebeca SoaresNo ratings yet
- Tecido MuscularDocument48 pagesTecido MuscularVAFNo ratings yet
- Aula 7 - Contração MuscularDocument25 pagesAula 7 - Contração MuscularJose GonsalvesNo ratings yet
- Tecido MuscularDocument6 pagesTecido MuscularLeo RamosNo ratings yet
- FisiologiaDocument9 pagesFisiologiakarol oliosiNo ratings yet
- FISIOLOGIA - RESUMO - Contração Do Musculo EsqueléticoDocument2 pagesFISIOLOGIA - RESUMO - Contração Do Musculo EsqueléticoLara Geovana BezerraNo ratings yet
- Centro Universitário Santo Agostinho Disciplina Organização Celular e Tecidual FarmáciaDocument8 pagesCentro Universitário Santo Agostinho Disciplina Organização Celular e Tecidual FarmáciaJOAO VICTOR COSTA LUSTOSANo ratings yet
- ARTIGO - Fisiologia Do Músculo EsqueléticoDocument32 pagesARTIGO - Fisiologia Do Músculo EsqueléticoKeyla SilvaNo ratings yet
- Tecido Muscular EsqueléticoDocument13 pagesTecido Muscular EsqueléticoMilene Borges MendesNo ratings yet
- Tecido Muscular - Departamento de Ciências Básicas - Unesp - Faculdade de Odontologia - Câmpus de AraçatubaDocument1 pageTecido Muscular - Departamento de Ciências Básicas - Unesp - Faculdade de Odontologia - Câmpus de AraçatubaDavi CastroNo ratings yet
- Aula 7 - Tecido MuscularDocument32 pagesAula 7 - Tecido MuscularBrenda SoutoNo ratings yet
- Apostila Fisiologia Neural Modelo 1Document17 pagesApostila Fisiologia Neural Modelo 1Alceu PaixãoNo ratings yet
- 07-01 Anatomia, Fisiologia Do Sistema CardiovascularDocument186 pages07-01 Anatomia, Fisiologia Do Sistema CardiovascularCarla CrivellaroNo ratings yet
- Apontamentos - Sebenta Fisiologia IDocument53 pagesApontamentos - Sebenta Fisiologia IJoao CarreiraNo ratings yet
- Atividade 3 de Projeto IntegradorDocument9 pagesAtividade 3 de Projeto Integradorvitor.vmNo ratings yet
- Slide Musculo LisoDocument25 pagesSlide Musculo LisoAdrya ThayanneNo ratings yet
- Apol 1 - Aspectos BiologicosDocument5 pagesApol 1 - Aspectos BiologicosJefersonmax DamataNo ratings yet
- Atividade de AprendizagemDocument4 pagesAtividade de AprendizagemVanessa CostaNo ratings yet
- Prova Medicina PSVO 2022-1Document12 pagesProva Medicina PSVO 2022-1jessica ynaieNo ratings yet
- Portifolio - HistologiaDocument22 pagesPortifolio - HistologiaAlex TauberNo ratings yet
- Sistema MuscularDocument16 pagesSistema MuscularRose LinonNo ratings yet
- Questões - HistologiaDocument25 pagesQuestões - HistologiaLopesMaxNo ratings yet
- Atlas de Miologia Veterinaria Cao e Equino Isabella RossiniDocument42 pagesAtlas de Miologia Veterinaria Cao e Equino Isabella RossiniLEANDRA TAVARES FURTADONo ratings yet
- Propriedades Da Carne FrescaDocument17 pagesPropriedades Da Carne Frescaromontezuma3607100% (1)
- Tecido MuscularDocument62 pagesTecido MuscularThiago Pimenta100% (1)
- Roteiro 1 - Tecido MuscularDocument9 pagesRoteiro 1 - Tecido MusculardievellenNo ratings yet
- Aula Teórica Histologia - Tecido Nervoso e MuscularDocument52 pagesAula Teórica Histologia - Tecido Nervoso e MuscularLeilane ArueiraNo ratings yet
- Tecidos RESUMODocument25 pagesTecidos RESUMOMariana RodriguesNo ratings yet
- FATORESQUEAFETAMAQUALIDADEDACARNEDocument11 pagesFATORESQUEAFETAMAQUALIDADEDACARNEMarilia C. PantaNo ratings yet
- 275 - Os BenefYcios Da Terapia Manual Na DesativaYYo Dos Pontos-GatilhoTCCDocument12 pages275 - Os BenefYcios Da Terapia Manual Na DesativaYYo Dos Pontos-GatilhoTCCAdriano Leao GomesNo ratings yet
- Apostila Dry Needling CursoDocument178 pagesApostila Dry Needling CursoKanandraNo ratings yet
- Perguntas de Prova HistologiaDocument7 pagesPerguntas de Prova HistologiaromuloNo ratings yet
- Respostas e Adaptações Metabólicas Ao ExercícioDocument8 pagesRespostas e Adaptações Metabólicas Ao ExercícioAndré CobeNo ratings yet
- Fatores Que Influenciam A Qualidade Da Carne BovinaDocument15 pagesFatores Que Influenciam A Qualidade Da Carne BovinaMichelle Simões de RezendeNo ratings yet
- Maturação Da Carne Bovina CompletoDocument16 pagesMaturação Da Carne Bovina CompletoJulianajsl100% (1)
- Aula 9 - Fisiologia Sensorial e MotoraDocument26 pagesAula 9 - Fisiologia Sensorial e MotoraValeskaSenaNo ratings yet
- Fisiologia Do Trabalho e Fisiopatologia Doencas OcupacionaisDocument112 pagesFisiologia Do Trabalho e Fisiopatologia Doencas OcupacionaisKaio RianNo ratings yet
- Função NeuromuscularDocument79 pagesFunção NeuromuscularClaudio Cruz SantosNo ratings yet
- Tecido MuscularDocument3 pagesTecido Muscular4pmpvz6hkrNo ratings yet
- Actina e Miosina Na Contração MuscularDocument19 pagesActina e Miosina Na Contração MuscularTiago SabençaNo ratings yet
- 6 Histologia AnimalDocument40 pages6 Histologia AnimalJoana GaiãoNo ratings yet