You might also like
- Clase 7, Hemorragia Del Primer TrimestreDocument7 pagesClase 7, Hemorragia Del Primer TrimestreCarlos Eduardo Machado AyalaNo ratings yet
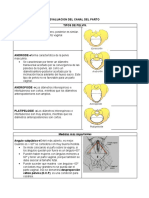
- Clase 3, Evaluacion Del Canal Del PartoDocument11 pagesClase 3, Evaluacion Del Canal Del PartoCarlos Eduardo Machado AyalaNo ratings yet
- Embarazo A Teěrmino 2Document10 pagesEmbarazo A Teěrmino 2Carlos Eduardo Machado AyalaNo ratings yet
- Clase 11, RPMDocument8 pagesClase 11, RPMCarlos Eduardo Machado AyalaNo ratings yet
- Clase 10, SX Parto PreterminoDocument7 pagesClase 10, SX Parto PreterminoCarlos Eduardo Machado AyalaNo ratings yet
- Clase 8, HUADocument8 pagesClase 8, HUACarlos Eduardo Machado AyalaNo ratings yet
- Clase 6, Embarazo ProlongadoDocument5 pagesClase 6, Embarazo ProlongadoCarlos Eduardo Machado AyalaNo ratings yet
- Duelo en Niños y AdolescentesDocument13 pagesDuelo en Niños y AdolescentesCarlos Eduardo Machado AyalaNo ratings yet
- DisfagiaDocument9 pagesDisfagiaCarlos Eduardo Machado AyalaNo ratings yet
- Clase 1, Generalidades ObstetriciaDocument6 pagesClase 1, Generalidades ObstetriciaCarlos Eduardo Machado AyalaNo ratings yet
- Clase 5, Control Prenatal - 1Document6 pagesClase 5, Control Prenatal - 1Carlos Eduardo Machado AyalaNo ratings yet
- Clase 5, Control Prenatal - 1Document6 pagesClase 5, Control Prenatal - 1Carlos Eduardo Machado AyalaNo ratings yet
- Clase 4 Induccion de Trabajo de PartoDocument10 pagesClase 4 Induccion de Trabajo de PartoCarlos Eduardo Machado AyalaNo ratings yet
- Enfoque de La HematuriaDocument6 pagesEnfoque de La HematuriaCarlos Eduardo Machado AyalaNo ratings yet
- Duelo en Niños y AdolescentesDocument13 pagesDuelo en Niños y AdolescentesCarlos Eduardo Machado AyalaNo ratings yet
- Secuencia Rapída de IntubacionDocument11 pagesSecuencia Rapída de IntubacionCarlos Eduardo Machado AyalaNo ratings yet
- Generalidades OrtopediaDocument12 pagesGeneralidades OrtopediaCarlos Eduardo Machado AyalaNo ratings yet
- Trauma de AbdomenDocument12 pagesTrauma de AbdomenCarlos Eduardo Machado AyalaNo ratings yet
- Pancreatitis AgudaDocument16 pagesPancreatitis AgudaCarlos Eduardo Machado AyalaNo ratings yet
- Infecciones Intraabdominales y Organos EspecificosDocument3 pagesInfecciones Intraabdominales y Organos EspecificosCarlos Eduardo Machado AyalaNo ratings yet
- Enfermedad Ulcerosa PépticaDocument10 pagesEnfermedad Ulcerosa PépticaCarlos Eduardo Machado AyalaNo ratings yet
- Enfermedad de CrohnDocument3 pagesEnfermedad de CrohnCarlos Eduardo Machado AyalaNo ratings yet
- Lineamientos 2018Document57 pagesLineamientos 2018yeisonNo ratings yet
- Guia - Rapida - Ahorro A La ManoDocument2 pagesGuia - Rapida - Ahorro A La ManoJuan PabloNo ratings yet
- PLAGIOCEFALIADocument3 pagesPLAGIOCEFALIACarlos Eduardo Machado AyalaNo ratings yet
- Infecciones IntraabdominalesDocument9 pagesInfecciones IntraabdominalesCarlos Eduardo Machado AyalaNo ratings yet
- Tejidos BlandosDocument8 pagesTejidos BlandosCarlos Eduardo Machado AyalaNo ratings yet
- Generalidades BacteriasDocument7 pagesGeneralidades BacteriasCarlos Eduardo Machado AyalaNo ratings yet
- 01 Plan de Área Transición 2019Document78 pages01 Plan de Área Transición 2019Carlos Eduardo Machado AyalaNo ratings yet
- MALTDocument3 pagesMALTCarlos Eduardo Machado AyalaNo ratings yet
- Evaluacion Final Personalidad JosephDocument8 pagesEvaluacion Final Personalidad JosephOscarSanmiguelNo ratings yet
- Cordova AndreaDocument109 pagesCordova AndreaRich RichNo ratings yet
- Derecho Penal Parte Especial-Tomo I-Tomas Galvez Villegas y Ricardo Rojas LeonDocument1,088 pagesDerecho Penal Parte Especial-Tomo I-Tomas Galvez Villegas y Ricardo Rojas Leonnomejoas enserio67% (3)
- Muelles Tarea 2Document11 pagesMuelles Tarea 2Marianita LisiNo ratings yet
- Politica de La Calidad y La Inocuidad de Los AlimentosDocument3 pagesPolitica de La Calidad y La Inocuidad de Los AlimentosJason CrossNo ratings yet
- 4 Practica - Calor Especifico - InformeDocument8 pages4 Practica - Calor Especifico - InformeEDER DANIEL ESCOBAR FLOREZNo ratings yet
- 54 Produccion Ecologica Cultivos Tropic AlesDocument76 pages54 Produccion Ecologica Cultivos Tropic AlesGuido Alex Luque HumpiriNo ratings yet
- Contactos Gays Pasivos Alcala Henares en MadridDocument2 pagesContactos Gays Pasivos Alcala Henares en MadridEroticoNo ratings yet
- Mpe - Semana 5 - Maraton 2023-IDocument22 pagesMpe - Semana 5 - Maraton 2023-IGonzalo AltamiranoNo ratings yet
- Ods 5 PDFDocument1 pageOds 5 PDFFabianoNo ratings yet
- Estudios de CasosDocument8 pagesEstudios de CasosLourdes X. MedinaNo ratings yet
- Diodo Zener Notas ElizabethDocument21 pagesDiodo Zener Notas ElizabethJohnNo ratings yet
- Apuntes de Materia y EnergiaDocument42 pagesApuntes de Materia y EnergiaSaul CalderonNo ratings yet
- Guia para La Educacion Inclusiva TeletonDocument15 pagesGuia para La Educacion Inclusiva TeletonVerónica Thomassiny AcostaNo ratings yet
- Taller #1 LácteosDocument3 pagesTaller #1 LácteosNataly NuñezNo ratings yet
- BIOQUIMICAS Bacillus ThuringiensisDocument11 pagesBIOQUIMICAS Bacillus ThuringiensisLewis Zúñiga DíazNo ratings yet
- Evaluación Práctica Jurídica II Meily Pérez CI 13.992.174Document28 pagesEvaluación Práctica Jurídica II Meily Pérez CI 13.992.174Meily PérezNo ratings yet
- Tarea 2 U2 Identificación de Normas Aplicables Equipo 6Document22 pagesTarea 2 U2 Identificación de Normas Aplicables Equipo 621460581No ratings yet
- Diapositivas Exposicion Campos MagneticosDocument15 pagesDiapositivas Exposicion Campos MagneticosMaria AcostaNo ratings yet
- La Influencia Del Ambiente FamiliarDocument2 pagesLa Influencia Del Ambiente Familiarosomoro0% (1)
- Plan de Tutoria. Aula 5toDocument7 pagesPlan de Tutoria. Aula 5toClarisa Herrera AcostaNo ratings yet
- Calculo Costos Produccion Litro Leche COLANTA.Document38 pagesCalculo Costos Produccion Litro Leche COLANTA.Pato Navarrete AvellánNo ratings yet
- Aspectos Culturales Que Influyen en La AlimentaciónDocument3 pagesAspectos Culturales Que Influyen en La AlimentaciónLuci CarmonaNo ratings yet
- Microbiologia F-7-9-2.Document8 pagesMicrobiologia F-7-9-2.Fabiana Reyes PrietoNo ratings yet
- RoyerDocument17 pagesRoyerMaicol Polo ArmasNo ratings yet
- Triptico VirreinatoDocument1 pageTriptico VirreinatoMaritza Salcedo MarquezNo ratings yet
- Clase IIIDocument53 pagesClase IIIALEXANDRA CASTAÑEDA BARRIOSNo ratings yet
- Ensayo - SIMCE 7mo BásicoDocument8 pagesEnsayo - SIMCE 7mo BásicoFrancisca Edwards RiuNo ratings yet
- 2do. Proyecto de AprendizajeDocument9 pages2do. Proyecto de AprendizajeArgenis AponteNo ratings yet
- InfografiasDocument3 pagesInfografiaslf249760No ratings yet