You might also like
- Libro 1Document2 pagesLibro 1Palomino AstridNo ratings yet
- No Metálicos IDocument4 pagesNo Metálicos IPalomino AstridNo ratings yet
- 3RA Evaluacion BioprocesosDocument5 pages3RA Evaluacion BioprocesosPalomino AstridNo ratings yet
- 3RA Evaluacion Bioprocesos PDFDocument5 pages3RA Evaluacion Bioprocesos PDFPalomino AstridNo ratings yet
- Biodiesel Con LipasasDocument3 pagesBiodiesel Con LipasasPalomino AstridNo ratings yet
- Simulación en Excel Del Comportamiento Del Cemento Portland Tipo I Mediante La Adición de 20Document14 pagesSimulación en Excel Del Comportamiento Del Cemento Portland Tipo I Mediante La Adición de 20Palomino AstridNo ratings yet
- Calder oDocument11 pagesCalder oPalomino AstridNo ratings yet
- Fenomenos EJEMPLO 1Document2 pagesFenomenos EJEMPLO 1Palomino AstridNo ratings yet
- Analisis Del Efecto de La Composiciopn Bioquimica de Los Principios Activos de Los Aceites Esenciales de RomeroDocument27 pagesAnalisis Del Efecto de La Composiciopn Bioquimica de Los Principios Activos de Los Aceites Esenciales de RomeroPalomino AstridNo ratings yet
- Refrigerador SolarDocument22 pagesRefrigerador SolarPalomino AstridNo ratings yet
- Analisis Del Efecto de La Composiciopn Bioquimica de Los Principios Activos de Los Aceites Esenciales de RomeroDocument27 pagesAnalisis Del Efecto de La Composiciopn Bioquimica de Los Principios Activos de Los Aceites Esenciales de RomeroPalomino AstridNo ratings yet
- Biodiesel Con LipasasDocument3 pagesBiodiesel Con LipasasPalomino AstridNo ratings yet
- Problemas Tarea 2 - PirometalurgiaDocument10 pagesProblemas Tarea 2 - PirometalurgiaPalomino AstridNo ratings yet
- Analisis Del Efecto de La Composiciopn Bioquimica de Los Principios Activos de Los Aceites Esenciales de RomeroDocument27 pagesAnalisis Del Efecto de La Composiciopn Bioquimica de Los Principios Activos de Los Aceites Esenciales de RomeroPalomino AstridNo ratings yet
- Calder oDocument11 pagesCalder oPalomino AstridNo ratings yet
- No Metálicos IDocument4 pagesNo Metálicos IPalomino AstridNo ratings yet
- Simulación del proceso de extracción de aceite esencial de molleDocument33 pagesSimulación del proceso de extracción de aceite esencial de mollePalomino Astrid100% (1)
- No Metálicos IDocument4 pagesNo Metálicos IPalomino AstridNo ratings yet
- MDocument33 pagesMPalomino AstridNo ratings yet
- Presentación 1Document3 pagesPresentación 1Palomino AstridNo ratings yet
- Escalamiento de Una Extractor de AceitesDocument14 pagesEscalamiento de Una Extractor de AceitesPalomino AstridNo ratings yet
- 1 y 2 de Diagrama de BloquesDocument4 pages1 y 2 de Diagrama de BloquesPalomino AstridNo ratings yet
- Ejercicio 2 Metodo de BiseccionDocument19 pagesEjercicio 2 Metodo de BiseccionPalomino AstridNo ratings yet
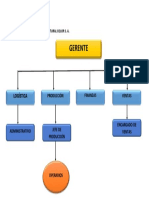
- OrganigramaDocument1 pageOrganigramaPalomino AstridNo ratings yet
- Cualidades de Un GerenteDocument13 pagesCualidades de Un GerentePalomino AstridNo ratings yet
- CuadroDocument6 pagesCuadroPalomino AstridNo ratings yet
- Plan Estratégico de Una EmpresaDocument42 pagesPlan Estratégico de Una EmpresaPalomino AstridNo ratings yet
- Gerencia IiDocument45 pagesGerencia IiPalomino AstridNo ratings yet
- Escalamiento de Una Planta de Extraccin de AceitesDocument1 pageEscalamiento de Una Planta de Extraccin de AceitesPalomino AstridNo ratings yet
- Plan Estratégico de Una EmpresaDocument42 pagesPlan Estratégico de Una EmpresaPalomino AstridNo ratings yet
- Ejes y PlanosDocument14 pagesEjes y PlanosbrauliolgNo ratings yet
- Fisiopatologia de La Preeclampsia 2015Document21 pagesFisiopatologia de La Preeclampsia 2015Lalo Angeles100% (1)
- Libro Frances HamerDocument3 pagesLibro Frances HamerPablo RivasNo ratings yet
- Articulo Petroleo Cura CancerDocument11 pagesArticulo Petroleo Cura CancerKolberKitasono100% (1)
- Uso de Drogas en El Personal de Enfermeria PDFDocument9 pagesUso de Drogas en El Personal de Enfermeria PDFFran QuirogaNo ratings yet
- Biorremediación de Suelos Contaminados Con Arsénico Mod.Document24 pagesBiorremediación de Suelos Contaminados Con Arsénico Mod.Henry Ccasa AlcaNo ratings yet
- Hipercolesterolemia Familiar: Causas, Síntomas y Tratamiento con EstatinasDocument10 pagesHipercolesterolemia Familiar: Causas, Síntomas y Tratamiento con EstatinasFelipe Javier Melis ArcosNo ratings yet
- Lectura 2017-6 ZelekDocument15 pagesLectura 2017-6 ZelekYolanda HernandezNo ratings yet
- Unidad IDocument318 pagesUnidad IMartin Penadillo DarwuinNo ratings yet
- Microbiologia AgricolaDocument36 pagesMicrobiologia AgricolaVALENTINA MONCAYO QUIGUANo ratings yet
- Catalogo de Piso PVC 2019 PDFDocument18 pagesCatalogo de Piso PVC 2019 PDFConstructora Emporio Dorado SASNo ratings yet
- CHR - Hansen-Cultivos para Quesos Continentales AmarillosDocument29 pagesCHR - Hansen-Cultivos para Quesos Continentales AmarillosEdgardo Josue Molina Martinez100% (1)
- Herencias genéticas y árboles genealógicosDocument8 pagesHerencias genéticas y árboles genealógicosHyllary BorjaNo ratings yet
- Matriz Iperc Vcn-Abengoa Rev10..Document34 pagesMatriz Iperc Vcn-Abengoa Rev10..Jose LuisNo ratings yet
- Valoración Basada en La Teoría de Kathryn EDocument18 pagesValoración Basada en La Teoría de Kathryn EMilagros DDíaz100% (1)
- Evaluación de impacto ambiental de la central hidroeléctrica de Tincoc mediante el método Batelle ColumbusDocument1 pageEvaluación de impacto ambiental de la central hidroeléctrica de Tincoc mediante el método Batelle ColumbusCesar Anthony Huaman ArceNo ratings yet
- Charla 5min JulioDocument22 pagesCharla 5min JulioDiego Molina ChavezNo ratings yet
- Proyecto Final CafeDocument41 pagesProyecto Final CafeVini MarcosNo ratings yet
- Revista El Hornero, Volumen 11, #5.Document105 pagesRevista El Hornero, Volumen 11, #5.Fernando Julio BioléNo ratings yet
- Manual Limpieza y SanitizaciónDocument8 pagesManual Limpieza y SanitizaciónLinoal NANo ratings yet
- Exposición AgroclimatologíaDocument15 pagesExposición AgroclimatologíaDaniel AlonsoNo ratings yet
- Estructura y función de los orgánulos celulares vegetalesDocument6 pagesEstructura y función de los orgánulos celulares vegetalesPerla Midary Dominguez CruzNo ratings yet
- Método PASS Teoría procesos cognitivosDocument28 pagesMétodo PASS Teoría procesos cognitivosjanaquevedoNo ratings yet
- 01 Urbanismo Antonio Ricaute - CANCHA Y PISTA DE TROTEDocument1 page01 Urbanismo Antonio Ricaute - CANCHA Y PISTA DE TROTEELIANANo ratings yet
- Aislamiento de PsicrofilosDocument3 pagesAislamiento de PsicrofilosSara Mucha BeraunNo ratings yet
- CARDIOLOGÍA - ResumenDocument29 pagesCARDIOLOGÍA - ResumenLucía GarcíaNo ratings yet
- 21 - FarmacoquimDocument9 pages21 - FarmacoquimMiguel Machaca Flores (QuimioFarma)No ratings yet
- Frotis vaginal y uretralDocument6 pagesFrotis vaginal y uretralCarlos Junior Polo PoloNo ratings yet
- Mitosis y MeiosisDocument2 pagesMitosis y MeiosisJaneane DavilaNo ratings yet
- Entrenamiento de La FuerzaDocument7 pagesEntrenamiento de La FuerzaCarlos GarcíaNo ratings yet