You might also like
- Ciclo Del Ácido TricarboxílicoDocument32 pagesCiclo Del Ácido TricarboxílicoDenissLopez100% (1)
- Acetil CoenzimaDocument3 pagesAcetil CoenzimaJose Dominguez100% (1)
- Presentación Nutrición Canina PDFDocument24 pagesPresentación Nutrición Canina PDFRocio Cabrera LoraNo ratings yet
- Qué Es La RumiaDocument2 pagesQué Es La RumiaFabiola Sanchez100% (1)
- Glucidos AvesDocument12 pagesGlucidos AvesCamila Tipaz100% (2)
- Oxidacion de Ácidos GrasosDocument1 pageOxidacion de Ácidos Grasosagustin zarate cruzNo ratings yet
- Exposicion Grupal - Metabolismo de Los Carbohidratos en Monogastricos CaninosDocument9 pagesExposicion Grupal - Metabolismo de Los Carbohidratos en Monogastricos CaninosAndrea GarcesNo ratings yet
- Metabolismo eritrocito glucosa ATP protege hemoglobinaDocument11 pagesMetabolismo eritrocito glucosa ATP protege hemoglobinaJGQM08No ratings yet
- Digestión de proteínas: Acción de enzimas pancreáticas sobre la caseínaDocument8 pagesDigestión de proteínas: Acción de enzimas pancreáticas sobre la caseínaCarlos CC100% (1)
- Cuestionario 11Document2 pagesCuestionario 11Mayker PanibraNo ratings yet
- Carbohidratos en Nutricion EquinaDocument5 pagesCarbohidratos en Nutricion EquinaAlex D' RoorshackNo ratings yet
- Rutas metabólicasDocument2 pagesRutas metabólicasAlvaro Hernández100% (1)
- Descarboxilacion OxidativaDocument34 pagesDescarboxilacion Oxidativaelvismartha50% (2)
- Estructura y funciones del tubo digestivoDocument21 pagesEstructura y funciones del tubo digestivoMónica Astete AnayaNo ratings yet
- Rutas MetabolicasDocument30 pagesRutas Metabolicasamilcar torresNo ratings yet
- Actividad en Clase de Cuerpos CetonicosDocument5 pagesActividad en Clase de Cuerpos CetonicosEzequiel SalazarNo ratings yet
- Ciclo de La UreaDocument6 pagesCiclo de La UreaBenaias Ramirez100% (1)
- Fosforilación Oxidativa Y Teoría QuimiosmóticaDocument35 pagesFosforilación Oxidativa Y Teoría QuimiosmóticaGema BernalNo ratings yet
- C5 Digestivo 1-RumiantesDocument101 pagesC5 Digestivo 1-RumiantesGerardo CF100% (1)
- 11 GluconeogénesissDocument13 pages11 GluconeogénesissEdmy Shasary Gutierrez Narvaez100% (2)
- COMPUESTOS RICOS EN ENERGÍA Y ENLACES RICOSDocument16 pagesCOMPUESTOS RICOS EN ENERGÍA Y ENLACES RICOSKarlita AlvarezNo ratings yet
- CETOGÉNESISDocument15 pagesCETOGÉNESISINick Salva PastorNo ratings yet
- Metabolismo de Otras HexosasDocument4 pagesMetabolismo de Otras HexosasMaria Zimbrelo Alcarria0% (1)
- Via de Las PentosasDocument25 pagesVia de Las PentosasHKlaudyNo ratings yet
- Metabolismo de Hemoglobina (DV)Document9 pagesMetabolismo de Hemoglobina (DV)Dary Deivy Varela AracheNo ratings yet
- Agar TioglicolatoDocument2 pagesAgar TioglicolatoAdrianaNo ratings yet
- Esquema CatabolismoDocument10 pagesEsquema CatabolismoBegoña FolNo ratings yet
- Jugos GastricosDocument8 pagesJugos GastricosBryan Giovanny ArmentaNo ratings yet
- Sindrome de La Vaca GordaDocument6 pagesSindrome de La Vaca GordaLea HimNo ratings yet
- Topografía Abdominal 1Document23 pagesTopografía Abdominal 1Jennifer RodriguezNo ratings yet
- Informe de Tracto Digestivo..Document6 pagesInforme de Tracto Digestivo..Javier Alejandro Vizcarra GonzalesNo ratings yet
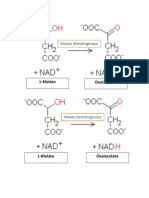
- La Enzima Malato DeshidrogenasaDocument2 pagesLa Enzima Malato DeshidrogenasaKaren Torres JuárezNo ratings yet
- Balantidium ColiDocument4 pagesBalantidium ColiWendy MoralesNo ratings yet
- Vías metabólicas de la glucosaDocument3 pagesVías metabólicas de la glucosaRebeca RioNo ratings yet
- Control hipotalámico de la reproducción a través de la KisspeptinaDocument191 pagesControl hipotalámico de la reproducción a través de la KisspeptinaBêtzzy GarcíaNo ratings yet
- 10 Alimentos Con Los 4 Aminoácidos LimitantesDocument2 pages10 Alimentos Con Los 4 Aminoácidos LimitantesDaraNolascoPalaciosNo ratings yet
- La Estructura de La InsulinaDocument3 pagesLa Estructura de La InsulinaAntonella Angulo Cruzado100% (1)
- El ciclo de la urea: transformación del amoníaco en ureaDocument65 pagesEl ciclo de la urea: transformación del amoníaco en ureaAlexandraMedinaCastilloNo ratings yet
- Ensayos QuimicosDocument5 pagesEnsayos QuimicosFrank BarreraNo ratings yet
- Los carbohidratos: fuente de energía y componentes estructuralesDocument4 pagesLos carbohidratos: fuente de energía y componentes estructuralesNicole AndradeNo ratings yet
- AGAR LIA 9.docxNNDocument2 pagesAGAR LIA 9.docxNNAndrea CohenNo ratings yet
- Vitamina E: 8 formas, funciones antioxidanteDocument12 pagesVitamina E: 8 formas, funciones antioxidanteKeiwin Gomez Villalobos0% (1)
- Ciclo de Cori: la circulación de glucosa y lactato entre músculo y hígadoDocument7 pagesCiclo de Cori: la circulación de glucosa y lactato entre músculo y hígadoJeanette PaulinaNo ratings yet
- Propiedades Químicas y Digestión Enzimática de Las ProteínasDocument5 pagesPropiedades Químicas y Digestión Enzimática de Las ProteínasNoriega VictoriaNo ratings yet
- METABOLISMO GLUCOGENODocument34 pagesMETABOLISMO GLUCOGENOEsmeralda CarreraNo ratings yet
- v9 Caso Clinico Feocromocitoma en Un PerroDocument5 pagesv9 Caso Clinico Feocromocitoma en Un PerroLiz AvalosNo ratings yet
- De Que Esta Compuesta La LenguaDocument3 pagesDe Que Esta Compuesta La LenguaMARGARITA HIDALGONo ratings yet
- Anatomía Ojo Pequeños AnimalesDocument35 pagesAnatomía Ojo Pequeños AnimalesNicole SaucedoNo ratings yet
- Metabolismo de La FructosaDocument9 pagesMetabolismo de La FructosaYenniffer Vicente Maya0% (1)
- Mascotas de Bolsillo. EnsayoDocument3 pagesMascotas de Bolsillo. EnsayoAndrea KuroNekoNo ratings yet
- DictiocaulosisDocument1 pageDictiocaulosisJosé López MartínezNo ratings yet
- Anatomía del aparato reproductor macho ovinoDocument6 pagesAnatomía del aparato reproductor macho ovinoRosita Vera Talles50% (2)
- Desarrollo embrionario de BocachicoDocument11 pagesDesarrollo embrionario de BocachicoPaula De Alba SurmayNo ratings yet
- Reacciones de La GlucolisisDocument5 pagesReacciones de La Glucolisissandy escorcia100% (1)
- Mecanismo de Acción de Las Toxinas de Bacillus AnthracisDocument2 pagesMecanismo de Acción de Las Toxinas de Bacillus AnthracisIzhmyy Dje100% (2)
- Cap 66 GuytonDocument7 pagesCap 66 GuytonRosario BarrientosNo ratings yet
- Practica #5.DETERMINACION DE LINFOCITOS T Y B.Document9 pagesPractica #5.DETERMINACION DE LINFOCITOS T Y B.JORGE EMILIANO RAMOS VILLALOBOS100% (1)
- DIURESISDocument2 pagesDIURESISJawetz100% (1)
- Taller GlucolisisDocument4 pagesTaller GlucolisisRoberto Jose Diaz CastroNo ratings yet
- RESÚMENESDocument5 pagesRESÚMENESMariel León BonifacioNo ratings yet
- La SDS PAGE-2D Más Que Una Técnica, "Un Método" A La Vanguardia en ProteómicaDocument21 pagesLa SDS PAGE-2D Más Que Una Técnica, "Un Método" A La Vanguardia en ProteómicaHernan Mauricio Rivera EscobarNo ratings yet
- Resultado Maestria UnalDocument1 pageResultado Maestria UnalHernan Mauricio Rivera EscobarNo ratings yet
- Manual de Practicas de Bioquimica BasicaDocument71 pagesManual de Practicas de Bioquimica BasicaHernan Mauricio Rivera EscobarNo ratings yet
- Circulares Mundo AventuraDocument3 pagesCirculares Mundo AventuraHernan Mauricio Rivera EscobarNo ratings yet
- Taller Bioquimica Cuarto PeriodoDocument2 pagesTaller Bioquimica Cuarto PeriodoHernan Mauricio Rivera Escobar80% (5)
- Transtornos de MenstruacionDocument11 pagesTranstornos de MenstruacionHernan Mauricio Rivera EscobarNo ratings yet
- Planeacion Total Ciencias JM 2010Document13 pagesPlaneacion Total Ciencias JM 2010Hernan Mauricio Rivera EscobarNo ratings yet
- Elaboracion de CervezaDocument6 pagesElaboracion de CervezaHernan Mauricio Rivera EscobarNo ratings yet
- Planeacion Quimica Once 2010Document10 pagesPlaneacion Quimica Once 2010Hernan Mauricio Rivera EscobarNo ratings yet
- Dia Del Ocio ProductivoDocument3 pagesDia Del Ocio ProductivoHernan Mauricio Rivera EscobarNo ratings yet
- Anexos Elaboracion de CervezaDocument3 pagesAnexos Elaboracion de CervezaHernan Mauricio Rivera EscobarNo ratings yet
- Codigo de EticaDocument4 pagesCodigo de EticaHernan Mauricio Rivera EscobarNo ratings yet
- Proyecto Del Manejo y Uso Del Tiempo LibreDocument7 pagesProyecto Del Manejo y Uso Del Tiempo LibreHernan Mauricio Rivera EscobarNo ratings yet
- Articulo Periodico Del Colegio VPHDocument3 pagesArticulo Periodico Del Colegio VPHHernan Mauricio Rivera EscobarNo ratings yet
- OLIMPIADAS Química 10º 2010Document3 pagesOLIMPIADAS Química 10º 2010Hernan Mauricio Rivera EscobarNo ratings yet
- Criterios de Evaluación y Promoción PropuestaDocument4 pagesCriterios de Evaluación y Promoción PropuestaHernan Mauricio Rivera EscobarNo ratings yet
- Energia Nuclear PresentacionDocument19 pagesEnergia Nuclear PresentacionHernan Mauricio Rivera EscobarNo ratings yet
- Planeacion Quimica 10 2010Document9 pagesPlaneacion Quimica 10 2010Hernan Mauricio Rivera EscobarNo ratings yet
- Programacion de Cursos Re Media Les y de Planes de Apoyo y Superacion 2010Document8 pagesProgramacion de Cursos Re Media Les y de Planes de Apoyo y Superacion 2010Hernan Mauricio Rivera EscobarNo ratings yet
- Guia de Soluciones Grado 10Document2 pagesGuia de Soluciones Grado 10Hernan Mauricio Rivera Escobar100% (1)
- Guia de Gases Grado 10Document3 pagesGuia de Gases Grado 10Hernan Mauricio Rivera EscobarNo ratings yet
- Actividad Vs Concentracion Enzimatica (1) LabDocument7 pagesActividad Vs Concentracion Enzimatica (1) LabHernan Mauricio Rivera Escobar50% (2)
- Informe Produccion de Cerveza Parte 2Document18 pagesInforme Produccion de Cerveza Parte 2Hernan Mauricio Rivera EscobarNo ratings yet
- OLIMPIADAS Química 11º 2010 LISTADocument3 pagesOLIMPIADAS Química 11º 2010 LISTAHernan Mauricio Rivera EscobarNo ratings yet
- Practica de Lab Modelos Moleculares e HibridacionDocument6 pagesPractica de Lab Modelos Moleculares e HibridacionHernan Mauricio Rivera Escobar100% (2)
- Formato de AutoevaluacionDocument2 pagesFormato de AutoevaluacionHernan Mauricio Rivera EscobarNo ratings yet
- Diagnostico 10º 2010 ListoDocument2 pagesDiagnostico 10º 2010 ListoHernan Mauricio Rivera EscobarNo ratings yet
- Informe Analitica Fluorometria LABDocument10 pagesInforme Analitica Fluorometria LABHernan Mauricio Rivera EscobarNo ratings yet
- Actividad de Aplicacion Anaglifos Lentes en Dos DimensionesDocument6 pagesActividad de Aplicacion Anaglifos Lentes en Dos DimensionesHernan Mauricio Rivera EscobarNo ratings yet
- Trabajo Practico de Quimica Organica Parte ADocument15 pagesTrabajo Practico de Quimica Organica Parte AjoanaNo ratings yet
- ATP: La molécula energética de la vidaDocument7 pagesATP: La molécula energética de la vidaMercedes Del Carmen Martinez ValverdeNo ratings yet
- Membrana PlasmaticaDocument27 pagesMembrana PlasmaticaIsa AguilarNo ratings yet
- Glosario de BioquimicaDocument5 pagesGlosario de Bioquimicaapi-257767200No ratings yet
- Activdad. Introducción CarbohidratosDocument2 pagesActivdad. Introducción CarbohidratosKelin Joselin Ceniceros SotoNo ratings yet
- Lipidos PDFDocument22 pagesLipidos PDFRozo MolinaNo ratings yet
- 3 A MonosacáridosDocument59 pages3 A MonosacáridosIvan HernándezNo ratings yet
- Principios de Ingeniería Metabólica: Biosíntesis de Ácidos Grasos y TriglicéridosDocument18 pagesPrincipios de Ingeniería Metabólica: Biosíntesis de Ácidos Grasos y TriglicéridosJessica RodríguezNo ratings yet
- 15 Preguntas Acerca de Las ProteinasDocument4 pages15 Preguntas Acerca de Las ProteinasNelsonArteagaTellez0% (1)
- Guia 05 Nomenclatura de Alcanos Alquenos y AlquinosDocument18 pagesGuia 05 Nomenclatura de Alcanos Alquenos y AlquinosPaula Pan PiragautaNo ratings yet
- Biomoléculas Orgánica 2022 CHS LIPIDOSDocument60 pagesBiomoléculas Orgánica 2022 CHS LIPIDOSSarai Hoyos VillavicencioNo ratings yet
- Metabolismo glucosa músculo esqueléticoDocument6 pagesMetabolismo glucosa músculo esqueléticoJuanPabloCorderoHenriquezNo ratings yet
- A10 McCabe Viscosidad en Líquidos (Nomográma)Document2 pagesA10 McCabe Viscosidad en Líquidos (Nomográma)PRizila Gomez RodriguezNo ratings yet
- Tabla ExcipientesDocument4 pagesTabla ExcipientesValeria GuerraNo ratings yet
- EsteroidesDocument16 pagesEsteroidesZu CRNo ratings yet
- Elementos de Quimica OrganicaDocument30 pagesElementos de Quimica Organicaceucih100% (1)
- Lípidos y LipoproteínasDocument44 pagesLípidos y LipoproteínasKarlos MartinezNo ratings yet
- CetonaDocument6 pagesCetonaSonia AyalaNo ratings yet
- Diapositiva BiologiaDocument1 pageDiapositiva Biologiaojedajohan432No ratings yet
- Informe Los LipidosDocument12 pagesInforme Los Lipidosnelcy aguilarNo ratings yet
- Fisiología HepáticaDocument23 pagesFisiología HepáticaBernabé LabonteNo ratings yet
- Cálculo fórmula láctea bebé 4 mesesDocument11 pagesCálculo fórmula láctea bebé 4 mesesDiego TrianaNo ratings yet
- Mapa Conceptual GlucidosDocument1 pageMapa Conceptual GlucidosHeidy CallejasNo ratings yet
- Informe 3 - Rasdsmlm PDFDocument18 pagesInforme 3 - Rasdsmlm PDFRAYDING DANIEL JULIO NARVAEZ ESTUDIANTE ACTIVONo ratings yet
- Guia QuimicaDocument6 pagesGuia Quimicajazmín cerratoNo ratings yet
- Ver21 Alfa RM1aDocument6 pagesVer21 Alfa RM1aJOSE DAVID PORRAS CORDOVANo ratings yet
- Hidrocarburos Ciclicos EjerciciosDocument4 pagesHidrocarburos Ciclicos EjerciciosValeria100% (2)
- 14 Absorción de Vitaminas 2021 1 9 EhhDocument13 pages14 Absorción de Vitaminas 2021 1 9 EhhYAMIL FABRICIO SOLDADO VALENCIANo ratings yet
- Plantilla de Excel para El Calculo de Dosis PediátricasDocument5 pagesPlantilla de Excel para El Calculo de Dosis PediátricaselaimygutierrezNo ratings yet
- Productos Naturales. AcetatosDocument58 pagesProductos Naturales. AcetatosAngela Sequera100% (1)