You might also like
- Program To Find The Area and Parameter of RectangleDocument51 pagesProgram To Find The Area and Parameter of RectangleShyam PrajaptiNo ratings yet
- IT SkillsDocument1 pageIT SkillsSrikanth KumarNo ratings yet
- Python Bootcamp Course OverviewDocument50 pagesPython Bootcamp Course OverviewIdo RaizbergNo ratings yet
- Ant Colony Optimization: A New Meta-HeuristicDocument8 pagesAnt Colony Optimization: A New Meta-Heuristicanil_invisibleNo ratings yet
- Solving Multiobjective Optimization Problems Using An Artificial Immune SystemDocument28 pagesSolving Multiobjective Optimization Problems Using An Artificial Immune SystemSebastián De La Cruz MartinezNo ratings yet
- Ant Colony Optimization For Continuous Domains: Krzysztof Socha, Marco DorigoDocument19 pagesAnt Colony Optimization For Continuous Domains: Krzysztof Socha, Marco DorigoJessica JaraNo ratings yet
- Bee ColonyDocument7 pagesBee ColonyLupu CiprianNo ratings yet
- Ant Colony Optimization BDocument8 pagesAnt Colony Optimization Bapi-3773589No ratings yet
- Ant Colony Optimization PDFDocument7 pagesAnt Colony Optimization PDFMati dell'ErbaNo ratings yet
- TSP Indin08Document7 pagesTSP Indin08GeorgehanyNo ratings yet
- Unit 4 MultimodelDocument10 pagesUnit 4 MultimodelYuvraj Singh RanaNo ratings yet
- Chaotic ElephantDocument4 pagesChaotic ElephantZainab SattarNo ratings yet
- A Hybrid Particle Swarm Evolutionary Algorithm For Constrained Multi-Objective Optimization Jingxuan WeiDocument18 pagesA Hybrid Particle Swarm Evolutionary Algorithm For Constrained Multi-Objective Optimization Jingxuan WeithavaselvanNo ratings yet
- ACO For Continuous and Mixed-Variable Optimization: Abstract. This Paper Presents How The Ant Colony Optimization (ACO)Document2 pagesACO For Continuous and Mixed-Variable Optimization: Abstract. This Paper Presents How The Ant Colony Optimization (ACO)Fatima LaqbitiNo ratings yet
- A Cellular Genetic Algorithm For Multiobjective OptimizationDocument12 pagesA Cellular Genetic Algorithm For Multiobjective Optimizationrogeriocosta37No ratings yet
- A Hybrid Ant Algorithm For The Airline CrewDocument11 pagesA Hybrid Ant Algorithm For The Airline CrewTatan BattagliaNo ratings yet
- Evolution Strategies With Additive Noise: A Convergence Rate Lower BoundDocument22 pagesEvolution Strategies With Additive Noise: A Convergence Rate Lower BoundHoldolf BetrixNo ratings yet
- ZzzChapter 3 - Section 3.3.8Document11 pagesZzzChapter 3 - Section 3.3.8Buddhi ChinthakaNo ratings yet
- Fuzzy Ant Colony Optimization For Optimal Control: Delft University of TechnologyDocument7 pagesFuzzy Ant Colony Optimization For Optimal Control: Delft University of TechnologyMed Cherif BellaouerNo ratings yet
- Overview of Algorithms For Swarm Intelligence PDFDocument14 pagesOverview of Algorithms For Swarm Intelligence PDFmenguemengueNo ratings yet
- Ant Colony Optimization1Document13 pagesAnt Colony Optimization1Dinda Tri PamungkasNo ratings yet
- Comparison of Genetic Algorithm and Particle Swarm OptimizationDocument6 pagesComparison of Genetic Algorithm and Particle Swarm OptimizationMikemekanicsNo ratings yet
- A Survey Paper On Solving Travelling Salesman Problem Using Bee Colony OptimizationDocument6 pagesA Survey Paper On Solving Travelling Salesman Problem Using Bee Colony Optimizationz1111234No ratings yet
- IEEE - Modified ACO Algorithm with Uniform Mutation for TSPDocument4 pagesIEEE - Modified ACO Algorithm with Uniform Mutation for TSPAniket SinghNo ratings yet
- Chaotic Bee Swarm Optimization Algorithm For Path Planning of Mobile RobotsDocument6 pagesChaotic Bee Swarm Optimization Algorithm For Path Planning of Mobile RobotsGeorgehanyNo ratings yet
- Co-Evolutionary Particle Swarm Optimization For Min-Max Problems Using Gaussian DistributionDocument6 pagesCo-Evolutionary Particle Swarm Optimization For Min-Max Problems Using Gaussian DistributionNaren PathakNo ratings yet
- Model For Quantifying RiskDocument12 pagesModel For Quantifying RiskIotalaseria PutuNo ratings yet
- Gecco2008 - OnctiDocument8 pagesGecco2008 - OnctiGustavo SánchezNo ratings yet
- Application of PSO, Artificial Bee Colony and Bacterial Foraging Optimization Algorithms To Economic Load Dispatch: An AnalysisDocument4 pagesApplication of PSO, Artificial Bee Colony and Bacterial Foraging Optimization Algorithms To Economic Load Dispatch: An AnalysispragatinareshNo ratings yet
- Evolutionary Multi-Objective Optimization in Materials ScienceDocument21 pagesEvolutionary Multi-Objective Optimization in Materials SciencevadivelbookNo ratings yet
- Dorado-Sevilla2021 Chapter AnInteractiveFrameworkToComparDocument16 pagesDorado-Sevilla2021 Chapter AnInteractiveFrameworkToComparZaki DahiaNo ratings yet
- Université Libre de Bruxelles: Tuning MAX-MIN Ant System With Off-Line and On-Line MethodsDocument23 pagesUniversité Libre de Bruxelles: Tuning MAX-MIN Ant System With Off-Line and On-Line Methodslcm3766lNo ratings yet
- Wong 2008Document6 pagesWong 2008باديس بن سديرةNo ratings yet
- Review of Ant Colony Optimization Algorithms Onvehicle Routing Problems and Introduction Toestimation-Based AcoDocument6 pagesReview of Ant Colony Optimization Algorithms Onvehicle Routing Problems and Introduction Toestimation-Based Acoadarsh_arya_1No ratings yet
- A Control Perspective For Centralized and Distributed Convex OptimizationDocument6 pagesA Control Perspective For Centralized and Distributed Convex Optimizationzxfeng168163.comNo ratings yet
- Ant Colony OptimizationDocument43 pagesAnt Colony OptimizationfreidyfriendsNo ratings yet
- Bee Colony Optimization With Local Search For TravDocument8 pagesBee Colony Optimization With Local Search For TravJamesNo ratings yet
- B. Tech, Batch, 2 Semester Subject Code: 18CS3064: Subject: Big Data OptimizationDocument11 pagesB. Tech, Batch, 2 Semester Subject Code: 18CS3064: Subject: Big Data Optimizationkrishnasai tadiboinaNo ratings yet
- A Parallel Ant Colony Optimization Algorithm With GPU-Acceleration Based On All-In-Roulette SelectionDocument5 pagesA Parallel Ant Colony Optimization Algorithm With GPU-Acceleration Based On All-In-Roulette SelectionJie FuNo ratings yet
- Firefly Paper1Document24 pagesFirefly Paper1vinod bhagatNo ratings yet
- Paper 15-Improving The Solution of Traveling Salesman Problem Using Genetic, Memetic Algorithm and Edge Assembly CrossoverDocument4 pagesPaper 15-Improving The Solution of Traveling Salesman Problem Using Genetic, Memetic Algorithm and Edge Assembly CrossoverEditor IJACSANo ratings yet
- Handling Constraints in Particle Swarm Optimization Using A Small Population SizeDocument11 pagesHandling Constraints in Particle Swarm Optimization Using A Small Population Sizedebasishmee5808No ratings yet
- Modularity Maximization in Networks by Variable Neighborhood SearchDocument14 pagesModularity Maximization in Networks by Variable Neighborhood SearchJohn ChoNo ratings yet
- LaumannsMarco Paper 246Document11 pagesLaumannsMarco Paper 246Toucha MayaNo ratings yet
- 2012 ANTS12 Chivilikhin UlyantsevDocument8 pages2012 ANTS12 Chivilikhin Ulyantsevyasir9909No ratings yet
- ShimuraDocument18 pagesShimuraDamodharan ChandranNo ratings yet
- Symmetry: An Improved Butterfly Optimization Algorithm For Engineering Design Problems Using The Cross-Entropy MethodDocument20 pagesSymmetry: An Improved Butterfly Optimization Algorithm For Engineering Design Problems Using The Cross-Entropy Methodomaima ghNo ratings yet
- A Particle Swarm Optimization For The Si PDFDocument15 pagesA Particle Swarm Optimization For The Si PDFRafael ValderramaNo ratings yet
- Ants 2002Document12 pagesAnts 2002api-3831656No ratings yet
- A Tutorial On Meta-Heuristics For OptimizationDocument36 pagesA Tutorial On Meta-Heuristics For Optimizationkrishnareddy_chintalaNo ratings yet
- Comparison of SVM, kNN, and Naive Bayes in Text ClassificationDocument10 pagesComparison of SVM, kNN, and Naive Bayes in Text ClassificationANDRES DAVID DOMINGUEZ ROZONo ratings yet
- Framework For Particle Swarm Optimization With Surrogate FunctionsDocument11 pagesFramework For Particle Swarm Optimization With Surrogate FunctionsLucas GallindoNo ratings yet
- Numerical Irreducible Decomposition Using PhcpackDocument20 pagesNumerical Irreducible Decomposition Using PhcpackNatasa BudisinNo ratings yet
- Applying Ants to Vehicle RoutingDocument12 pagesApplying Ants to Vehicle RoutingMuhammad AriefNo ratings yet
- Non-Dominated Archiving Multi-Colony Ant Algorithm For Multi-Objective Optimization - Application ToDocument14 pagesNon-Dominated Archiving Multi-Colony Ant Algorithm For Multi-Objective Optimization - Application ToKarim EL BouyahyiouyNo ratings yet
- Bare Demo of IEEEtran Cls For IEEE ConferencesDocument10 pagesBare Demo of IEEEtran Cls For IEEE Conferencesup8975479No ratings yet
- Exploiting_Problem_Decomposition_in_Multi-objectivDocument16 pagesExploiting_Problem_Decomposition_in_Multi-objectiva.frank.liceNo ratings yet
- Differential Evolution With Ensemble of ConstraintDocument9 pagesDifferential Evolution With Ensemble of ConstraintMBADJOUN DanielNo ratings yet
- The Optimal Design and Simulation of Helical Spring Based On Particle Swarm Algorithm and MatlabDocument10 pagesThe Optimal Design and Simulation of Helical Spring Based On Particle Swarm Algorithm and MatlabvenkiteshksNo ratings yet
- 10.1007@s10898 007 9256 8Document18 pages10.1007@s10898 007 9256 8NikosNo ratings yet
- Reynoso Meza2011Document6 pagesReynoso Meza2011Ankit PrajapatiNo ratings yet
- PracticeDocument4 pagesPracticeAnoop ChauhanNo ratings yet
- Resiliencepatterns 141105153710 Conversion Gate02Document81 pagesResiliencepatterns 141105153710 Conversion Gate02Dhiraj ThakurNo ratings yet
- GOOSE Configuration ExampleDocument20 pagesGOOSE Configuration ExampleANTONIO SOLISNo ratings yet
- Lingo 18 Users Manual (001-500)Document500 pagesLingo 18 Users Manual (001-500)Horacio Jesus Prado GarciaNo ratings yet
- C Program of Round Robin FCFS and SJF CPU SchedulingDocument4 pagesC Program of Round Robin FCFS and SJF CPU SchedulingAman RajNo ratings yet
- Partial Exam QuestionsDocument19 pagesPartial Exam QuestionsfaurandradaNo ratings yet
- UNIX LINUX Training Course, Best UNIX LINUX Training Institute Hyderabad, India - NareshITDocument5 pagesUNIX LINUX Training Course, Best UNIX LINUX Training Institute Hyderabad, India - NareshITAnand Dhanalakota0% (1)
- SiVarc Multiuser PDFDocument51 pagesSiVarc Multiuser PDFseyed ebrahim hosseiniNo ratings yet
- Facebook ThriftDocument13 pagesFacebook ThriftYashwant Prakash LamboleNo ratings yet
- LightGBM - An In-Depth Guide PythonDocument26 pagesLightGBM - An In-Depth Guide PythonRadha KilariNo ratings yet
- Introduction to References: Understanding Types, Objects, Variables, Pointers and ReferencesDocument16 pagesIntroduction to References: Understanding Types, Objects, Variables, Pointers and ReferencesJeremy PhoNo ratings yet
- Generics and Collections OverviewDocument134 pagesGenerics and Collections OverviewDeepa SubramanyamNo ratings yet
- Bluetooth - Programming Microsoft Windows CE .NET - 3rd Edition - Douglas BolingDocument35 pagesBluetooth - Programming Microsoft Windows CE .NET - 3rd Edition - Douglas Bolingjei liNo ratings yet
- Database Management System Lab Prelim Q1Document7 pagesDatabase Management System Lab Prelim Q1Michelle EsperalNo ratings yet
- Learning Guide-Dcit 55-Advanced Database SystemDocument5 pagesLearning Guide-Dcit 55-Advanced Database SystemJenjie Villaseñor Dela PeñaNo ratings yet
- COMP3711: Design and Analysis of Algorithms: Tutorial 5 HkustDocument31 pagesCOMP3711: Design and Analysis of Algorithms: Tutorial 5 Hkustfsvvdfhf100% (1)
- CSS Final Practical QBDocument17 pagesCSS Final Practical QB13TYCM-IOm DerleNo ratings yet
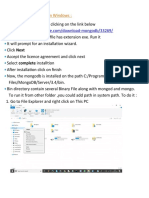
- How to Install MongoDB on Windows in 10 Easy StepsDocument20 pagesHow to Install MongoDB on Windows in 10 Easy Stepsvikas yadavNo ratings yet
- Chapter-01 Programming LanguagesDocument10 pagesChapter-01 Programming LanguagesSyed Mir Talha ZobaedNo ratings yet
- DA-IICT BTech Graduate Kishan Thesiya's ResumeDocument1 pageDA-IICT BTech Graduate Kishan Thesiya's ResumeAmit Shukla 20037No ratings yet
- PL Class 01 02022013Document33 pagesPL Class 01 02022013Adil AhmadNo ratings yet
- Quine McCluskey MethodDocument19 pagesQuine McCluskey MethodNathaly Lizbeth Sipiran MorilloNo ratings yet
- BTech in AI CurriculumDocument7 pagesBTech in AI CurriculumCnp SethupathiNo ratings yet
- BSC (Hons) Web TechnologiesDocument17 pagesBSC (Hons) Web TechnologiesRoubin DhallapahNo ratings yet
- Notes - Assembler Directives PDFDocument23 pagesNotes - Assembler Directives PDFAshritha G100% (1)
- DBMS NotesDocument18 pagesDBMS NotesYamanNo ratings yet
- Unit-4 NetworkingDocument47 pagesUnit-4 NetworkingShashikant MoreNo ratings yet