You might also like
- Mitosis and MeiosisDocument13 pagesMitosis and Meiosis420192420192No ratings yet
- Sordaria Lab ReportDocument12 pagesSordaria Lab Reportapi-305324400100% (1)
- Sordaria Genetics Lab ReportDocument11 pagesSordaria Genetics Lab Reportapi-305324573100% (1)
- Sordaria Lab ReportDocument9 pagesSordaria Lab Reportapi-218590219100% (1)
- Sordaria Lab ReportDocument4 pagesSordaria Lab Reportapi-253660813No ratings yet
- Sordaria Lab Report FinalDocument12 pagesSordaria Lab Report Finalapi-316516464100% (1)
- Sordaria Lab ReportDocument9 pagesSordaria Lab Reportapi-217409009No ratings yet
- On The Morphology of The Chromosome Group in Brachystola MagnaDocument18 pagesOn The Morphology of The Chromosome Group in Brachystola MagnaOya KaptanoğluNo ratings yet
- D. melanogaster Productivity Increased with Banana and Pear EnrichmentDocument6 pagesD. melanogaster Productivity Increased with Banana and Pear EnrichmentEdgar A. ÇolmenárezNo ratings yet
- Determining Plant Species Dominance Using Terrestrial Sampling TechniquesDocument9 pagesDetermining Plant Species Dominance Using Terrestrial Sampling TechniquesJohn Jerald VillamancaNo ratings yet
- Staining TechniquesDocument19 pagesStaining TechniquesSwayamprakash PatelNo ratings yet
- BIOL 240 Lab Report 1Document11 pagesBIOL 240 Lab Report 1Ben CharlesNo ratings yet
- Lab Report For Micro 210 AJH1Document11 pagesLab Report For Micro 210 AJH1ahathaw17No ratings yet
- The Neurohistological TechniquesDocument4 pagesThe Neurohistological Techniquesapi-3846255100% (2)
- Extrachromosomal Inheritance: Kappa Particles in ParameciumDocument3 pagesExtrachromosomal Inheritance: Kappa Particles in ParameciumJoshua NabablitNo ratings yet
- Stool Examination (4)Document29 pagesStool Examination (4)محمد رحيم حسن محمودNo ratings yet
- Breeding The Papaya - Carica Papaya PDFDocument11 pagesBreeding The Papaya - Carica Papaya PDFchikkannaswamyNo ratings yet
- Chromosome MappingDocument22 pagesChromosome MappingVipin100% (1)
- Analyzing The PV92 Locus On Chromosome 16 For The Alu Insertion Through The Polymerase Chain Reaction and Agarose Gel ElectrophoresisDocument10 pagesAnalyzing The PV92 Locus On Chromosome 16 For The Alu Insertion Through The Polymerase Chain Reaction and Agarose Gel ElectrophoresisJohansen C. Pico100% (1)
- TranscriptionDocument72 pagesTranscriptionMurthy MandalikaNo ratings yet
- Cell Counting Using A HemacytometerDocument3 pagesCell Counting Using A HemacytometerKris Cahyo Mulyatno100% (1)
- Polytene and Lampbrush ChromosomesDocument3 pagesPolytene and Lampbrush Chromosomesneeru.bhagatNo ratings yet
- BandingDocument3 pagesBandingBacaro JosyndaNo ratings yet
- Albugo: White Rust of CrucifersDocument34 pagesAlbugo: White Rust of CrucifersMuhammad MushtaqNo ratings yet
- Spectrophotometric Estimation of Escitalopram OxalateDocument3 pagesSpectrophotometric Estimation of Escitalopram Oxalateapi-19918842No ratings yet
- Arthropod Diversity Lab ReportDocument3 pagesArthropod Diversity Lab ReportJudge Michael GarzaNo ratings yet
- Life Cycle of OedogoniumDocument16 pagesLife Cycle of OedogoniumMuskan Sachdeva 0047No ratings yet
- Isolation and Enumeration of Bacteria in Water and FoodDocument30 pagesIsolation and Enumeration of Bacteria in Water and FoodOld Lake100% (1)
- QuantifyingBiodiversity - Spider LabDocument4 pagesQuantifyingBiodiversity - Spider LabJohn OsborneNo ratings yet
- Karyotype Analysis in Zea Mays L. Var. Everta (Popcorn) Cultivated Within Owerri, Southeast NigeriaDocument6 pagesKaryotype Analysis in Zea Mays L. Var. Everta (Popcorn) Cultivated Within Owerri, Southeast NigeriaImpact JournalsNo ratings yet
- Methods To Study Soil Microbial DiversityDocument25 pagesMethods To Study Soil Microbial Diversityrd2165scribdNo ratings yet
- M - 23 Polytene ChromosomeDocument3 pagesM - 23 Polytene ChromosomeDr. Tapan Kr. DuttaNo ratings yet
- In Vitro Pollen Germinationin Pigeonpes 2013Document5 pagesIn Vitro Pollen Germinationin Pigeonpes 2013Sabesan TNo ratings yet
- Lab ProtocolDocument5 pagesLab Protocolanmol3No ratings yet
- Podocarpus: The Largest Genus of ConifersDocument9 pagesPodocarpus: The Largest Genus of ConifersFouzia Youseph100% (1)
- Detection of Nucleic AcidDocument13 pagesDetection of Nucleic AcidAfrah AlatasNo ratings yet
- Comparing DNA Sequences To Understand Evolutionary Relationships With BlastDocument3 pagesComparing DNA Sequences To Understand Evolutionary Relationships With Blastflyawayxx13No ratings yet
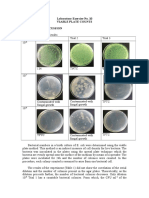
- Laboratory Exercise No. 10 Viable Plate Counts Results and DiscussionDocument3 pagesLaboratory Exercise No. 10 Viable Plate Counts Results and Discussionvanessa olga100% (1)
- Cytogenetic Slide PreparationDocument9 pagesCytogenetic Slide PreparationRicardo PalmaNo ratings yet
- BOTANY First - PAPER (Algae, Lichens and Bryophyta)Document3 pagesBOTANY First - PAPER (Algae, Lichens and Bryophyta)GuruKPO0% (1)
- Fate Mapping: Understanding Embryonic DevelopmentDocument27 pagesFate Mapping: Understanding Embryonic DevelopmentSaad SakalNo ratings yet
- Coriander Seed Yield VarietiesDocument10 pagesCoriander Seed Yield VarietiesVikki Nandeshwar100% (1)
- Embryonic Fate MapDocument25 pagesEmbryonic Fate MapImran MohsinNo ratings yet
- Molecular PhylogeneticsDocument4 pagesMolecular PhylogeneticsAb WassayNo ratings yet
- C. asiaticum leaves promote wound healingDocument5 pagesC. asiaticum leaves promote wound healingxiuhtlaltzinNo ratings yet
- MeiosisDocument8 pagesMeiosisJohn Chiyu Garde-Labite Azuki100% (1)
- Experiment 4Document5 pagesExperiment 4enieynazNo ratings yet
- Lab Gen. Physio 11Document7 pagesLab Gen. Physio 11Eugene Lucino CodisNo ratings yet
- Quelch Tree Lab ReportDocument9 pagesQuelch Tree Lab ReporttquelchNo ratings yet
- Ap Cellular Respiration LabDocument8 pagesAp Cellular Respiration Labapi-382372564No ratings yet
- Quiz 3 AnswersDocument3 pagesQuiz 3 AnswersEdilberto PerezNo ratings yet
- Chromosomal Aberrations: Deficiency or DeletionDocument4 pagesChromosomal Aberrations: Deficiency or Deletionvandv printsNo ratings yet
- Temperature Gradient Gel Electrophoresis-SenDocument7 pagesTemperature Gradient Gel Electrophoresis-Sensivagurunathan.P100% (1)
- Chapter 11 - Meristem Culture For Virus Free Plants - 2013 - Plant Tissue Cultur PDFDocument8 pagesChapter 11 - Meristem Culture For Virus Free Plants - 2013 - Plant Tissue Cultur PDFLuis BermudezNo ratings yet
- 9.2 Water Transport - Exam Questions: Root HairDocument4 pages9.2 Water Transport - Exam Questions: Root HairShaeNo ratings yet
- Recombinant ReportDocument4 pagesRecombinant Report门门No ratings yet
- Hyperthermophiles - MicrobiologyDocument15 pagesHyperthermophiles - MicrobiologysathishNo ratings yet
- APA Unknown Lab ReportDocument9 pagesAPA Unknown Lab ReportMiss DNo ratings yet
- Meiosis PaperDocument7 pagesMeiosis Paperawilson338No ratings yet
- ( ) Unit5 Meiosis MCQDocument11 pages( ) Unit5 Meiosis MCQEric ChangNo ratings yet
- Science: Quarter 4 - Module 1 The Human Digestive SystemDocument43 pagesScience: Quarter 4 - Module 1 The Human Digestive SystemGenesis SarengoNo ratings yet
- Biology F4 Review Summary NoteDocument117 pagesBiology F4 Review Summary NoteSocdal AbdiNo ratings yet
- Common Mistakes-BiologyDocument8 pagesCommon Mistakes-BiologyteahockNo ratings yet
- Gene Linkage & Mapping Using Tetrad Analysis in The Fungus Sordaria Fimicola 1-GlaseDocument24 pagesGene Linkage & Mapping Using Tetrad Analysis in The Fungus Sordaria Fimicola 1-GlaseCHRISTEROPNo ratings yet
- Biology Today and Tomorrow With Physiology 5th Edition Starr Test BankDocument24 pagesBiology Today and Tomorrow With Physiology 5th Edition Starr Test Bankdariusadele6ncs100% (33)
- Cell DivisionDocument72 pagesCell Divisionnor ainiNo ratings yet
- Meiosis With Pipe CleanersDocument2 pagesMeiosis With Pipe Cleanersapi-3004364130% (1)
- Meiosis and Gamteogenesis - 1 PDFDocument447 pagesMeiosis and Gamteogenesis - 1 PDFJOSE FRANCISCO FRANCO NAVIANo ratings yet
- Unit 3 BiologyDocument183 pagesUnit 3 BiologyBeverly Crawford75% (8)
- Biology Practical Part 2Document23 pagesBiology Practical Part 2jiya singh100% (2)
- Group 1Document31 pagesGroup 1John DanaoNo ratings yet
- General Mcqs PDFDocument5 pagesGeneral Mcqs PDFJunaid ahmad lucky sahaa100% (1)
- Kami Export - Cristina Cotra - Meiosis ActivityDocument7 pagesKami Export - Cristina Cotra - Meiosis ActivityCristina CotraNo ratings yet
- KVPY 2013 SA SolutionDocument23 pagesKVPY 2013 SA SolutionArshKhan50% (2)
- Cell CycleDocument12 pagesCell CycleVega, Charles Gabriel G.No ratings yet
- GenBio SMILE LP4 Q1 WEEK4Document16 pagesGenBio SMILE LP4 Q1 WEEK4bobby carsonNo ratings yet
- Variation F 5Document8 pagesVariation F 5Angie Kong Su MeiNo ratings yet
- Biology (Form 4)Document25 pagesBiology (Form 4)Aloysius RajNo ratings yet
- Principles of Inheritance and Variations Questions With AnswersDocument8 pagesPrinciples of Inheritance and Variations Questions With AnswersPrajwal dNo ratings yet
- The Cell Cycle Activity 1Document5 pagesThe Cell Cycle Activity 1sugarxglossNo ratings yet
- Anaphase II The Sister Chromatids Are Pulled Apart and HeadDocument20 pagesAnaphase II The Sister Chromatids Are Pulled Apart and Headain mikanNo ratings yet
- Biology The Essentials 1st Edition Hoefnagels Test BankDocument40 pagesBiology The Essentials 1st Edition Hoefnagels Test Bankblanchetoj0fg100% (24)
- Questions On MeiosisDocument10 pagesQuestions On MeiosisChelain BoucherNo ratings yet
- Ncert Biology Chapter I0Document11 pagesNcert Biology Chapter I0sai arunNo ratings yet
- Life Sciences Easy To Score p2 Marking Guideline (O - 230909 - 144945Document17 pagesLife Sciences Easy To Score p2 Marking Guideline (O - 230909 - 144945Silindile PreciousNo ratings yet
- Cell Division Study NotesDocument9 pagesCell Division Study NotesShaira Galban GamponNo ratings yet
- Botany Outline Characteristics of Living Things and Manifestations of LifeDocument33 pagesBotany Outline Characteristics of Living Things and Manifestations of LifeElma GonzalesNo ratings yet
- Mitosis and MeiosisDocument4 pagesMitosis and MeiosisMicah Porcal ArelladoNo ratings yet
- Cell Division Class 9Document4 pagesCell Division Class 9pillapandurangapatruduNo ratings yet