You might also like
- EnzymeKinetics by P.C. Misra Professor, Department of Biochemistry Lucknow University, Lucknow-226 007Document21 pagesEnzymeKinetics by P.C. Misra Professor, Department of Biochemistry Lucknow University, Lucknow-226 007Dr. SHIVA AITHALNo ratings yet
- Microbiology of Water and Waste Water ManagementDocument27 pagesMicrobiology of Water and Waste Water ManagementDr. SHIVA AITHAL100% (2)
- 50 Books For ChildrenDocument32 pages50 Books For ChildrenDr. SHIVA AITHALNo ratings yet
- FOOD MICROBIOLOGY Spoilage Poisoning and PreservationDocument13 pagesFOOD MICROBIOLOGY Spoilage Poisoning and PreservationDr. SHIVA AITHAL100% (1)
- Hanuman Chalisa English MeaningDocument3 pagesHanuman Chalisa English MeaningSaurabh SaxenaNo ratings yet
- Glycolysis and TCA Cycle - ShuttlesDocument1 pageGlycolysis and TCA Cycle - ShuttlesDr. SHIVA AITHALNo ratings yet
- Air Microbiology 2009Document13 pagesAir Microbiology 2009Dr. SHIVA AITHAL100% (9)
- Visit To ISDN BayalaluDocument16 pagesVisit To ISDN BayalaluDr. SHIVA AITHAL100% (2)
- Glycolysis and TCA Cycle Enzymes and EC Reference NumbersDocument1 pageGlycolysis and TCA Cycle Enzymes and EC Reference NumbersDr. SHIVA AITHALNo ratings yet
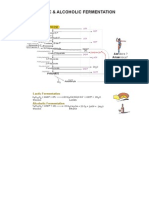
- Lactic and Alcoholic FermentationDocument1 pageLactic and Alcoholic FermentationDr. SHIVA AITHALNo ratings yet
- GlycolysisDocument1 pageGlycolysisDr. SHIVA AITHALNo ratings yet
- Structure of DNA by CrickDocument8 pagesStructure of DNA by CrickDr. SHIVA AITHALNo ratings yet
- Modelo Doble Helice Watson y CrickDocument2 pagesModelo Doble Helice Watson y Crickangelferp100% (2)
- Micro Teaching Skills ComponentsDocument35 pagesMicro Teaching Skills ComponentsDr. SHIVA AITHAL89% (221)
- 3G Wireless NetworksDocument16 pages3G Wireless NetworksAli QureshiNo ratings yet
- A Brief Foray Into The World of Nano Materials by Sarbari BhattacharyaDocument17 pagesA Brief Foray Into The World of Nano Materials by Sarbari BhattacharyaDr. SHIVA AITHALNo ratings yet
- Management of Libraries and Information Sciences by Naznin BanuDocument12 pagesManagement of Libraries and Information Sciences by Naznin BanuDr. SHIVA AITHALNo ratings yet
- Preamble To The Constitution of India by Prema NagaleDocument12 pagesPreamble To The Constitution of India by Prema NagaleDr. SHIVA AITHAL100% (1)
- Biotechnology in Agriculture Development by KS ManjunathDocument15 pagesBiotechnology in Agriculture Development by KS ManjunathDr. SHIVA AITHAL100% (1)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Chapter 16 Lippincott BiochemistryDocument57 pagesChapter 16 Lippincott BiochemistryMeysam SajjadiNo ratings yet
- Q2 W1 W2 W3 General Biology 1Document8 pagesQ2 W1 W2 W3 General Biology 1Samantha ManibogNo ratings yet
- General Introduction and Basic Biochemical PrinciplesDocument6 pagesGeneral Introduction and Basic Biochemical PrinciplescelecosibNo ratings yet
- Exam 4 Study GuideDocument4 pagesExam 4 Study Guideca2aokereNo ratings yet
- Test 1 - Aerobic RespirationDocument3 pagesTest 1 - Aerobic Respirationhasbiallah1306No ratings yet
- 2ND Activity Frezel EjemparDocument5 pages2ND Activity Frezel EjemparRONALD ARTILLERONo ratings yet
- 2.glycolysis & Oxidation of PyruvateDocument12 pages2.glycolysis & Oxidation of Pyruvateقتيبه خالد دحام خلفNo ratings yet
- Bioenergetics Photosynthesis and Energy FlowDocument4 pagesBioenergetics Photosynthesis and Energy Flowvin ocangNo ratings yet
- AP Biology Outline - Chapter 7Document4 pagesAP Biology Outline - Chapter 7Omar LopezNo ratings yet
- UNIT - II: Lipid MetabolismDocument27 pagesUNIT - II: Lipid Metabolismgaurav saxena100% (1)
- Photosynthesis Hhmi BiointeractiveDocument7 pagesPhotosynthesis Hhmi BiointeractivemariaNo ratings yet
- 26 Biology 2-16-08 Cellular RespirationDocument48 pages26 Biology 2-16-08 Cellular Respirationpotato macchiatoNo ratings yet
- Cie A2 - 12Document18 pagesCie A2 - 12S Ms.No ratings yet
- ABG Answers Sheet - 2Document1 pageABG Answers Sheet - 2khryss100% (1)
- 2) Reaksi Biosintesis Metabolit SekunderDocument22 pages2) Reaksi Biosintesis Metabolit SekunderAndiaisyah macorawalieNo ratings yet
- Preparing For An ExamDocument2 pagesPreparing For An ExamHamdan FatahNo ratings yet
- GluconeogenesisDocument3 pagesGluconeogenesisLinhNguyeNo ratings yet
- Photosynthesis (KEY)Document6 pagesPhotosynthesis (KEY)kylevNo ratings yet
- Nutrition in Plants Ppt'sDocument67 pagesNutrition in Plants Ppt's5 D 160300 MANASVI REDDY CHITTEDINo ratings yet
- Ureacycle ConversionDocument31 pagesUreacycle Conversionarun231187No ratings yet
- Photosynthesis MIND Maps by Om MishraDocument3 pagesPhotosynthesis MIND Maps by Om MishrasandhyaatiwariNo ratings yet
- Cytoplasm and OrganellesDocument22 pagesCytoplasm and OrganellesKrishiaDeVeraNo ratings yet
- Bioenergetics 1Document30 pagesBioenergetics 1suni alditaNo ratings yet
- BIOENERGETICS Trans - IncDocument3 pagesBIOENERGETICS Trans - IncChino Paolo SamsonNo ratings yet
- Calculation of Substrate Oxidation Rates in Vivo From Gaseous ExchangeDocument7 pagesCalculation of Substrate Oxidation Rates in Vivo From Gaseous ExchangeRodrigo CastilloNo ratings yet
- Biochemical ParametersDocument39 pagesBiochemical ParametersdystoNo ratings yet
- Cyclic PhotophosphorylationDocument10 pagesCyclic PhotophosphorylationJenard Atos33% (3)
- بايو بيبر 1 د.جميلة السمهريDocument8 pagesبايو بيبر 1 د.جميلة السمهريOzgan SüleymanNo ratings yet
- Bio Lesson 21 RespiDocument48 pagesBio Lesson 21 RespiChlzy ArdienteNo ratings yet
- Biochimica MetabolicaDocument195 pagesBiochimica MetabolicaGianluca100% (1)