You might also like
- Biogénesis mitocondrial y ejercicio: señalización por calcio y ATPDocument14 pagesBiogénesis mitocondrial y ejercicio: señalización por calcio y ATPMari CapiNo ratings yet
- Copia de Proyecto ABI 1.1Document16 pagesCopia de Proyecto ABI 1.1Dante Huitron HernandezNo ratings yet
- Fisiología Del Trabajo de PartoDocument9 pagesFisiología Del Trabajo de PartogalileofigaromlNo ratings yet
- Capitulo XviDocument16 pagesCapitulo XviIvette FloresNo ratings yet
- Segundos MensajerosDocument6 pagesSegundos MensajerosDARI LUZ ARIAS MORENONo ratings yet
- Adaptaciones moleculares al entrenamiento aeróbicoDocument32 pagesAdaptaciones moleculares al entrenamiento aeróbicoRODRIGO CACHON VILLAMEDIANANo ratings yet
- Vías de Transducción Intracelular de SeñalesDocument8 pagesVías de Transducción Intracelular de SeñalesCristina PazmiñoNo ratings yet
- Examen Final 19-9-2019Document2 pagesExamen Final 19-9-2019Joseph ReyesNo ratings yet
- Mecanismos Moleculares en La Adaptacion Al EjercicioDocument8 pagesMecanismos Moleculares en La Adaptacion Al EjercicioJULIA LUNA GONZALEZNo ratings yet
- F. Adaptacion MolecularDocument36 pagesF. Adaptacion Molecularjavier patricio cando casanovaNo ratings yet
- Factores Que Intervienen en La Contracción MuscularDocument3 pagesFactores Que Intervienen en La Contracción MuscularJuan Norris AldacoNo ratings yet
- Final Bioquimica FMED UBA 9-2-18Document3 pagesFinal Bioquimica FMED UBA 9-2-18Cande AmiconeNo ratings yet
- FarmacodinamiaDocument29 pagesFarmacodinamiaMercedes RamosNo ratings yet
- Daryenel Huitron Autocoides y Segundos MensajerosDocument22 pagesDaryenel Huitron Autocoides y Segundos MensajerosDaryenel HuitrónNo ratings yet
- Roig, Jorge - Intimidades de La AMPK MuscularDocument5 pagesRoig, Jorge - Intimidades de La AMPK Musculardiegus D masNo ratings yet
- AMPK Regulador Metabolico MaestroDocument9 pagesAMPK Regulador Metabolico MaestroBrajan LeónNo ratings yet
- Acidosis Metabólica Inducida Por El EjercicioDocument5 pagesAcidosis Metabólica Inducida Por El EjercicioAndrea villadiegoNo ratings yet
- Sobre El MagnesioDocument10 pagesSobre El MagnesioFranko LopezNo ratings yet
- AMPK-mTOR en hipertrofia, atrofia y autofagiaDocument3 pagesAMPK-mTOR en hipertrofia, atrofia y autofagiaJavier YañezNo ratings yet
- La fosfocreatina como depósito de energía muscularDocument11 pagesLa fosfocreatina como depósito de energía muscularJackelyne AlejosNo ratings yet
- Bases Energéticas Del Ejercicio en El Caballo Contraccion MuscularDocument29 pagesBases Energéticas Del Ejercicio en El Caballo Contraccion MuscularAngel Chavez100% (1)
- El AMP BiologiaDocument4 pagesEl AMP BiologiaJuan Guillermo Cañaveral TabaresNo ratings yet
- 14 Senales CelularesDocument57 pages14 Senales CelularesElbio AlvarezNo ratings yet
- Rutas de SeñalizaciónDocument7 pagesRutas de SeñalizaciónFlor de MariaNo ratings yet
- GlucolisisDocument5 pagesGlucolisisCintya Mabel Pinaya CruzNo ratings yet
- Actividad Obligatoria - Ud3Document3 pagesActividad Obligatoria - Ud3RoNo ratings yet
- Fisiologia de La EreccionDocument8 pagesFisiologia de La EreccionAlejo W. GomezNo ratings yet
- Cap 036Document4 pagesCap 036Luis Felipe GuzmánNo ratings yet
- Cárnicos - Libro TraducidoDocument20 pagesCárnicos - Libro TraducidoRicardo AvilaNo ratings yet
- Calcio Articulo VertebradosDocument11 pagesCalcio Articulo VertebradosYar CBNo ratings yet
- Preguntes FISIODocument8 pagesPreguntes FISIOmargotNo ratings yet
- Señalización Fosfolipasa CDocument4 pagesSeñalización Fosfolipasa CVictor Vitoria83% (6)
- Ciclo Del MagnesioDocument10 pagesCiclo Del MagnesioSandra Lizbeth100% (1)
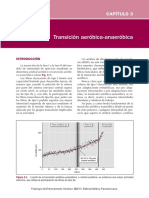
- Transición Aeróbica-AnaeróbicaDocument9 pagesTransición Aeróbica-AnaeróbicaJessica Izchel Velasco BarajasNo ratings yet
- Mecanismo de Acción de Los MineralesDocument5 pagesMecanismo de Acción de Los MineralesErikaParedesSulcaNo ratings yet
- El Papel Funcional de La Calcineurina en La Hipertrofia EspañolDocument10 pagesEl Papel Funcional de La Calcineurina en La Hipertrofia Españoledison0mej0asNo ratings yet
- Mapas Conceptuales de FisiopatologiaDocument13 pagesMapas Conceptuales de FisiopatologiaKely SrosNo ratings yet
- Receptores CelularesDocument5 pagesReceptores CelularesPamela Alejandra Marín VillalónNo ratings yet
- Lopez Chicharro AerobicoDocument9 pagesLopez Chicharro AerobicoPilar Bravo Salas100% (1)
- Receptores, Segundos Mensajeros y Vias de Senalizacion1Document14 pagesReceptores, Segundos Mensajeros y Vias de Senalizacion1SantiagoNo ratings yet
- 5. Receptores, Segundos Mensajeros y Vias de Senalizacion1Document14 pages5. Receptores, Segundos Mensajeros y Vias de Senalizacion1esrn4laboratorioNo ratings yet
- Atp y Vias de Señalizacion CelularDocument4 pagesAtp y Vias de Señalizacion CelularMarq TintaNo ratings yet
- 2023 - P1 - TEMA A RespuestasDocument2 pages2023 - P1 - TEMA A RespuestasvaalenmediNo ratings yet
- Clase 2012 10 SenalesDocument46 pagesClase 2012 10 Senalespau_sebNo ratings yet
- 3 MusculoDocument4 pages3 MusculoNitxia GuzmanNo ratings yet
- Unidad 3 Metabolismo, Catabolismo y AnabolismoDocument17 pagesUnidad 3 Metabolismo, Catabolismo y Anabolismolizeth gaambaNo ratings yet
- CALCITONINADocument1 pageCALCITONINAGeranni RondonNo ratings yet
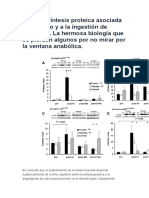
- mTOR y Síntesis Proteica Asociada Al Ejercicio yDocument5 pagesmTOR y Síntesis Proteica Asociada Al Ejercicio yRuben Congosto SanchezNo ratings yet
- Cambios Bioquimicos Del PescadoDocument63 pagesCambios Bioquimicos Del PescadoAngela Valencia Chavez50% (2)
- 11 Contraccion MuscularDocument54 pages11 Contraccion MuscularDiego NolascoNo ratings yet
- AMPK y Los Cambios Metabólicos Después Del Ejercicio - IMPRESODocument6 pagesAMPK y Los Cambios Metabólicos Después Del Ejercicio - IMPRESOJuani CantellanosNo ratings yet
- Acoplamineto Excitación - ContracciónDocument4 pagesAcoplamineto Excitación - ContracciónKim Hyun JoongNo ratings yet
- Tema 8. Respuestas Del Organismo Ante La Actividad FisicaDocument19 pagesTema 8. Respuestas Del Organismo Ante La Actividad Fisicajulin400No ratings yet
- 2023 - P1 - TEMA C RespuestasDocument2 pages2023 - P1 - TEMA C RespuestasvaalenmediNo ratings yet
- Lesión CelularDocument11 pagesLesión CelularMaria Fernanda Carvajal CastillaNo ratings yet
- Ayudas ergogénicas y nutricionalesFrom EverandAyudas ergogénicas y nutricionalesRating: 5 out of 5 stars5/5 (2)
- La Ciencia Del Metabolismo: Metabolismo sin rodeosFrom EverandLa Ciencia Del Metabolismo: Metabolismo sin rodeosRating: 5 out of 5 stars5/5 (1)
- 25 Consejos para Aumentar La Seguridad en Linux - Noticias - CSIDocument8 pages25 Consejos para Aumentar La Seguridad en Linux - Noticias - CSIНаталля ПавіненNo ratings yet
- Planificador de ClasesDocument51 pagesPlanificador de ClasesRodrigo Meneses100% (1)
- 01 Aprender A Jugar Al Ajedrez - IntroducciónDocument9 pages01 Aprender A Jugar Al Ajedrez - Introducciónlunalo100% (1)
- Asamblea de conejos protege a sus crías de los cazadoresDocument2 pagesAsamblea de conejos protege a sus crías de los cazadoresAndrea Carrasco SaezNo ratings yet
- Coleccion Cuentos KawesqarDocument128 pagesColeccion Cuentos KawesqarНаталля ПавіненNo ratings yet
- Imtroduccion Al Metodo ABNDocument23 pagesImtroduccion Al Metodo ABNanon_533992282100% (2)
- La CreatividadDocument1 pageLa CreatividadНаталля ПавіненNo ratings yet
- Prueba Los Niños de La Cruz Del Sur 7°BDocument3 pagesPrueba Los Niños de La Cruz Del Sur 7°BНаталля ПавіненNo ratings yet
- Guia Estudio PUEBLOS 15 NovDocument1 pageGuia Estudio PUEBLOS 15 NovНаталля ПавіненNo ratings yet
- Conservando La Flora y Fauna Amenazada - CONAFDocument152 pagesConservando La Flora y Fauna Amenazada - CONAFCeyanezNo ratings yet
- Article - La Sexualidad Infantil y La Autorregulación - Casilda RodrigañezDocument3 pagesArticle - La Sexualidad Infantil y La Autorregulación - Casilda RodrigañezDani Sanchez Diaz100% (1)
- Curso de Linux para Novatos Brutos y Extremadamente Torpes (WWW - Yovani.netne - Net)Document303 pagesCurso de Linux para Novatos Brutos y Extremadamente Torpes (WWW - Yovani.netne - Net)Geovanni100% (1)
- M. Fukuoka - La Revolución de Un Rastrojo (Brizna de Paja) PDFDocument96 pagesM. Fukuoka - La Revolución de Un Rastrojo (Brizna de Paja) PDFBe GreenNo ratings yet
- Telar Patagon Guia PrincipiantesDocument155 pagesTelar Patagon Guia PrincipiantesFrancisca Alvial Vargas100% (3)
- 200 Labores Ganchillo OrkideaDocument124 pages200 Labores Ganchillo OrkideaMyrna Soledad Quintana100% (9)
- Ficha171 Metodo MatteDocument10 pagesFicha171 Metodo MatteHeidi Cuevas Venegas100% (1)
- Manual de Construccion en Tierra - Gernot MinkeDocument222 pagesManual de Construccion en Tierra - Gernot Minkestrempler100% (24)
- M. Fukuoka - La Revolución de Un Rastrojo (Brizna de Paja) PDFDocument96 pagesM. Fukuoka - La Revolución de Un Rastrojo (Brizna de Paja) PDFBe GreenNo ratings yet
- BioquimicaDocument1 pageBioquimicaНаталля ПавіненNo ratings yet
- M. Fukuoka - La Revolución de Un Rastrojo (Brizna de Paja) PDFDocument96 pagesM. Fukuoka - La Revolución de Un Rastrojo (Brizna de Paja) PDFBe GreenNo ratings yet
- Air CrackDocument8 pagesAir CrackLeira Nahitsirc IhcamilNo ratings yet
- VeganismoDocument28 pagesVeganismoWilder CorreaNo ratings yet
- Monografia DesnutriciónDocument18 pagesMonografia DesnutriciónguillermoperNo ratings yet
- Especies de Nitrógeno Reactivo RNSDocument13 pagesEspecies de Nitrógeno Reactivo RNSRubi Lopez EspadaNo ratings yet
- Proyecto de Biologia 2do de Bachillerato Parcial 2Document18 pagesProyecto de Biologia 2do de Bachillerato Parcial 2Yury RamirezNo ratings yet
- Adelgazar y Realzar La BellezaDocument89 pagesAdelgazar y Realzar La BellezafedejomedinaNo ratings yet
- Alimentos ricos en proteínasDocument10 pagesAlimentos ricos en proteínasluis100% (1)
- Formato Plan de Clase Periodo 16Document7 pagesFormato Plan de Clase Periodo 16DALIA ORTEGANo ratings yet
- El caldo primigenio y el origen de la vidaDocument4 pagesEl caldo primigenio y el origen de la vidaVictor Flores VasquezNo ratings yet
- Guía Genética Molecular Noveno (3) 27Document5 pagesGuía Genética Molecular Noveno (3) 27valeria valentina novoa paredesNo ratings yet
- Introducción Al CitoesqueletoDocument5 pagesIntroducción Al CitoesqueletoAina cuixart gimoNo ratings yet
- Memorias 2016Document189 pagesMemorias 2016Gera CorralesNo ratings yet
- GD - Biologia-pENSAR 3Document16 pagesGD - Biologia-pENSAR 3Alejandra Masgoret CuellarNo ratings yet
- 038 D QuinuaDocument28 pages038 D QuinuaJoe Huaranccay QuispeNo ratings yet
- Extraccion de GelatinaDocument12 pagesExtraccion de GelatinaIng Max de la CruzNo ratings yet
- Cta4 U5 Sesion 01geneticaDocument4 pagesCta4 U5 Sesion 01geneticarosa luzNo ratings yet
- Manual B Forrajeros 04Document5 pagesManual B Forrajeros 04cuastumalitoNo ratings yet
- Clase 15 - Aislamiento de Adn A Partir de Celulas VegetalesDocument4 pagesClase 15 - Aislamiento de Adn A Partir de Celulas Vegetalesyuzumagloria3No ratings yet
- Repaso BiologiaDocument42 pagesRepaso BiologiaCosmic latteNo ratings yet
- Preguntas Explicadas BiologiaDocument62 pagesPreguntas Explicadas BiologiaBrandon AfanadorNo ratings yet
- Caracterizacion Estructural de ProteinasDocument9 pagesCaracterizacion Estructural de ProteinasMarta FerliniNo ratings yet
- Alimentación Del DeportistaDocument5 pagesAlimentación Del DeportistaJulio MontielNo ratings yet
- Sesiones de Cts MartinezDocument9 pagesSesiones de Cts MartinezEdgar Fernando Martinez Sanchez50% (2)
- Nutri LiteDocument51 pagesNutri LiteRaul IzquierdoNo ratings yet
- Cómo comer como un fisicoculturista profesionalDocument5 pagesCómo comer como un fisicoculturista profesionalhorfeoNo ratings yet
- Plan de Negocios en Pro y Comer. de Quinua OrganicaDocument40 pagesPlan de Negocios en Pro y Comer. de Quinua OrganicaNikolai Solis Loaiza100% (1)
- Propiedades Funcionales de Las Proteinas de La LecheDocument21 pagesPropiedades Funcionales de Las Proteinas de La LecheCarMarquzNo ratings yet
- BiomoléculasDocument2 pagesBiomoléculasAdrian RodriguezNo ratings yet
- CXS - 072s NORMA PARA PREPARADOS PARA LACTANTES Y PREPARADOSDocument19 pagesCXS - 072s NORMA PARA PREPARADOS PARA LACTANTES Y PREPARADOSxacvierNo ratings yet
- GloriaDocument8 pagesGloriaHelencitaPaucarNo ratings yet
- Primer SemestreDocument74 pagesPrimer SemestreJuank Torres AriasNo ratings yet