You might also like
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Pharmacology - Antibiotics Flash CardsDocument20 pagesPharmacology - Antibiotics Flash CardsJamil100% (2)
- Pelvic Inflammatory DiseaseDocument9 pagesPelvic Inflammatory DiseaseSohanInduwaraGamage100% (2)
- Peds Drug Mixing PDFDocument18 pagesPeds Drug Mixing PDFeialcaheNo ratings yet
- Introduction To MicrobiologyDocument52 pagesIntroduction To MicrobiologyHoward Barlomento100% (3)
- Antibiotic Resistance and Extended Spectrum Beta-Lactamases: Types, Epidemiology and TreatmentDocument35 pagesAntibiotic Resistance and Extended Spectrum Beta-Lactamases: Types, Epidemiology and TreatmentRoby Aditya SuryaNo ratings yet
- Project Neem SanjayDocument40 pagesProject Neem Sanjaysinghjsanjay885No ratings yet
- Environmental Hazards and Human Health Study GuideDocument5 pagesEnvironmental Hazards and Human Health Study GuidezsiddiquiNo ratings yet
- OREGANO Oil of Oregano Rivals Modern Antibiotic DrugsDocument1 pageOREGANO Oil of Oregano Rivals Modern Antibiotic DrugsDonald-Virginia KjellbergNo ratings yet
- Scientific Critique of EvolutionDocument19 pagesScientific Critique of Evolutionasilvestre19807No ratings yet
- Isolation of Bacterial Pathogens From Eye Infection Patients From QuettaDocument9 pagesIsolation of Bacterial Pathogens From Eye Infection Patients From QuettaEngr Faisal Saleem JanjuaNo ratings yet
- CellulitisDocument8 pagesCellulitisTricia Llorin100% (1)
- What's The Best Antibiotic For A Staph Infection - Treatments and RisksDocument3 pagesWhat's The Best Antibiotic For A Staph Infection - Treatments and RisksSundaramoorthy SelvanathanNo ratings yet
- Cefadroxil (Cephalosporin Generasi I)Document5 pagesCefadroxil (Cephalosporin Generasi I)Yustia SariNo ratings yet
- Government of Nepal Has Banned The Folowing Medicines For Production, Sale-Distribution and Import (Oral and Parenteral Use) On 2040/3/13Document7 pagesGovernment of Nepal Has Banned The Folowing Medicines For Production, Sale-Distribution and Import (Oral and Parenteral Use) On 2040/3/13Jitendra YadavNo ratings yet
- Norfloxillin 200 Sol.: Bi D VW WJB 200 MJDocument2 pagesNorfloxillin 200 Sol.: Bi D VW WJB 200 MJSharfina Akter BithiNo ratings yet
- Biology of Nutrition in Growing AnimalsDocument599 pagesBiology of Nutrition in Growing AnimalsSam Rafael Bravo GutierrezNo ratings yet
- Meningitis Pediatrics in Review Dic 2015Document15 pagesMeningitis Pediatrics in Review Dic 2015Edwin VargasNo ratings yet
- Febrile Neutropenia GuidelineDocument8 pagesFebrile Neutropenia GuidelineAslesa Wangpathi PagehgiriNo ratings yet
- Time and ConcentrationDocument2 pagesTime and ConcentrationikrimakhaerunnisaNo ratings yet
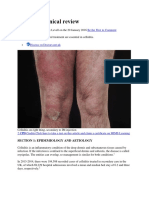
- Cellulitis: Clinical Review: Be The First To CommentDocument9 pagesCellulitis: Clinical Review: Be The First To CommentAnonymous 1nMTZWmzNo ratings yet
- SDSU BIO 350 Exam3 2010 VerA (With Answers)Document5 pagesSDSU BIO 350 Exam3 2010 VerA (With Answers)Ernie RoweNo ratings yet
- Ace InhibitorsDocument15 pagesAce InhibitorsCarolyn Conn EdwardsNo ratings yet
- Spektrum AntibakteriDocument1 pageSpektrum AntibakteriZulfikar Hizbul IslamiNo ratings yet
- Previous HSE Questions From The Chapter "CHEMISTRY IN EVERY DAY LIFE"Document2 pagesPrevious HSE Questions From The Chapter "CHEMISTRY IN EVERY DAY LIFE"Nikhil MathewNo ratings yet
- Bala Resume New FormatDocument4 pagesBala Resume New FormateldinhenryNo ratings yet
- AntibigramDocument27 pagesAntibigramManal A.FatahNo ratings yet
- Introduction To ChemotherapyDocument28 pagesIntroduction To ChemotherapyShivsharan B. DhaddeNo ratings yet
- JMPR - 15 August, 2012 IssueDocument127 pagesJMPR - 15 August, 2012 IssuetoronyoraNo ratings yet
- Argumentative Research EssayDocument5 pagesArgumentative Research Essayckjennings6100% (1)
- Assessment of Antibiotic Utilization in Sepsis Order Sets Initiated in The Emergency Department Draft 2Document45 pagesAssessment of Antibiotic Utilization in Sepsis Order Sets Initiated in The Emergency Department Draft 2api-652562554No ratings yet