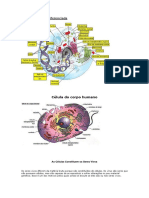
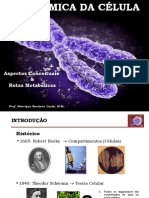
You might also like
- Apostila MaqFluxo EG PDFDocument109 pagesApostila MaqFluxo EG PDFÍthalo SantanaNo ratings yet
- Rotulagem Ciclo de VidaDocument40 pagesRotulagem Ciclo de VidaEduardo Sá Freire de Souza100% (1)
- Auditoria AmbientalDocument12 pagesAuditoria AmbientalEduardo Sá Freire de SouzaNo ratings yet
- Rio +20 Aula 2Document1 pageRio +20 Aula 2Eduardo Sá Freire de SouzaNo ratings yet
- 3186-Texto Do Artigo-12931-1-10-20150929Document22 pages3186-Texto Do Artigo-12931-1-10-20150929Eduardo Sá Freire de SouzaNo ratings yet
- Ensaios Padrão de Areia de FundiçãoDocument3 pagesEnsaios Padrão de Areia de FundiçãoEduardo Sá Freire de SouzaNo ratings yet
- Parte I - Introducao Aos Processos de Fabricao MetalicaDocument66 pagesParte I - Introducao Aos Processos de Fabricao MetalicaAnonymous 9rJe2lOskxNo ratings yet
- Treino ABCDocument1 pageTreino ABCEduardo Sá Freire de SouzaNo ratings yet
- Catalogo ContinentalDocument88 pagesCatalogo ContinentalEduardo Sá Freire de SouzaNo ratings yet
- Tabela de Folgas JQC PDFDocument1 pageTabela de Folgas JQC PDFEduardo Sá Freire de SouzaNo ratings yet
- Adm MotivaçãoDocument3 pagesAdm MotivaçãoEduardo Sá Freire de SouzaNo ratings yet
- Teorias de MotivaçãoDocument6 pagesTeorias de MotivaçãoEduardo Sá Freire de SouzaNo ratings yet
- Auditoria AmbientalDocument12 pagesAuditoria AmbientalEduardo Sá Freire de SouzaNo ratings yet
- Tramitacao-VTS 1 CCJC - PEC 241-2016Document9 pagesTramitacao-VTS 1 CCJC - PEC 241-2016Eduardo Sá Freire de SouzaNo ratings yet
- Resumo Formulario-CalcVetDocument4 pagesResumo Formulario-CalcVetEduardo Sá Freire de SouzaNo ratings yet
- Apostila MODocument16 pagesApostila MOEduardo Sá Freire de SouzaNo ratings yet
- Gesta o Ambient AlDocument4 pagesGesta o Ambient AlEduardo Sá Freire de SouzaNo ratings yet
- Análise de TensõesDocument3 pagesAnálise de TensõesEduardo Sá Freire de SouzaNo ratings yet
- Terça-Feira, 16 de Agosto de 2011Document1 pageTerça-Feira, 16 de Agosto de 2011Eduardo Sá Freire de SouzaNo ratings yet
- Resistência Dos Materiais I - ResumoDocument22 pagesResistência Dos Materiais I - ResumoEduardo Sá Freire de SouzaNo ratings yet
- Desbloquear o Home TheaterDocument1 pageDesbloquear o Home TheaterEduardo Sá Freire de SouzaNo ratings yet
- De Que Arvore Voce CaiuDocument24 pagesDe Que Arvore Voce CaiuAcacio NetoNo ratings yet
- Tema 7 - Transcrição Do Dna - Versão Final PDFDocument50 pagesTema 7 - Transcrição Do Dna - Versão Final PDFPedro SilvaNo ratings yet
- Artigo (Consumo de Álcool e Hipertrofia)Document20 pagesArtigo (Consumo de Álcool e Hipertrofia)Vinicius RiannNo ratings yet
- RESUMO Metabolismo Das LipoproteínasDocument7 pagesRESUMO Metabolismo Das LipoproteínasSalomão SandesNo ratings yet
- Perguntas e Respostas PDFDocument14 pagesPerguntas e Respostas PDFAnaCristinaCorreiaNo ratings yet
- Aula 6 Regulação Da Expressão GênicaDocument30 pagesAula 6 Regulação Da Expressão GênicaDiego Balbine100% (1)
- Bioquímica - Resumo 2017Document82 pagesBioquímica - Resumo 2017Joana AquinoNo ratings yet
- Via Das Pentoses FosfatoDocument24 pagesVia Das Pentoses FosfatoCesar SchmollerNo ratings yet
- Biologia e Bioquimica HumanaDocument5 pagesBiologia e Bioquimica HumanaHenrique CamposNo ratings yet
- Célula Humana IndiferenciadaDocument180 pagesCélula Humana IndiferenciadaAna Paula Pereira Cunha100% (1)
- Correção F3Document8 pagesCorreção F3Ana BaptistaNo ratings yet
- Lista Exercícios - Bioquímica MetabólicaDocument2 pagesLista Exercícios - Bioquímica MetabólicaLucas PintanelNo ratings yet
- MEDRESUMOS 2016 - IMUNOLOGIA 16 - Imunologia Dos TumoresDocument7 pagesMEDRESUMOS 2016 - IMUNOLOGIA 16 - Imunologia Dos TumoresneudodmNo ratings yet
- Síntese ProteicaDocument32 pagesSíntese ProteicaCunha PrOMasterNo ratings yet
- Questionário I - ProcariotosDocument4 pagesQuestionário I - ProcariotosGiovana Álice de Souza SantosNo ratings yet
- Aula4.1 em PDF - Especializações de SuperfícieDocument27 pagesAula4.1 em PDF - Especializações de SuperfícieSabrine CamargoNo ratings yet
- Aula 2 Bioquimica Da Transformacao Do Músculo Esquelético em Carne. Parte IDocument19 pagesAula 2 Bioquimica Da Transformacao Do Músculo Esquelético em Carne. Parte IRodrigo AcioliNo ratings yet
- Aula 9 - Diferenciação Celular e Morte CelularDocument10 pagesAula 9 - Diferenciação Celular e Morte CelularAmanda HodelNo ratings yet
- Gliconeogênese (Modo de Compatibilidade)Document16 pagesGliconeogênese (Modo de Compatibilidade)Guto Braga100% (1)
- Aula 02 - Bioquímica Da CélulaDocument27 pagesAula 02 - Bioquímica Da CélulaRenato ViniciusNo ratings yet
- EXERCÍCIOS SOBRE MITOSE, MEIOSE E GAMETOGÊNESE (Respondido)Document2 pagesEXERCÍCIOS SOBRE MITOSE, MEIOSE E GAMETOGÊNESE (Respondido)Vinicius Evangelista da SilvaNo ratings yet
- Aula 1 - Morfologia e Estrutura CelularDocument37 pagesAula 1 - Morfologia e Estrutura CelularhathexselNo ratings yet
- Bases Citológicas Da HerançaDocument27 pagesBases Citológicas Da HerançaLúcio Eduardo FerreiraNo ratings yet
- PATOLOGIADocument1 pagePATOLOGIAmonagisaNo ratings yet
- 4000 QUESTÕES DE BIOLOGIA-Prof. Leandro GomesDocument362 pages4000 QUESTÕES DE BIOLOGIA-Prof. Leandro GomesAna Beatriz Almeida100% (9)
- Slide de PatologiaDocument38 pagesSlide de PatologiaWilber LeonidasNo ratings yet
- Estrutura Do NeurónioDocument12 pagesEstrutura Do Neuróniodavid joaoNo ratings yet
- Ficha de Leitura - OdtDocument15 pagesFicha de Leitura - OdtHanustúrio Jorge cossaNo ratings yet
- Botanica - Osmose Difusao e PlasmoliseDocument33 pagesBotanica - Osmose Difusao e PlasmolisejanainalopesmarinNo ratings yet
- IM Aula 2 ProteinasDocument11 pagesIM Aula 2 ProteinasTarcísio CarterNo ratings yet
- Aula 1 - Célula VegetalDocument33 pagesAula 1 - Célula VegetalJoab Ana MenezesNo ratings yet