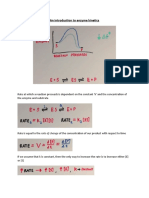
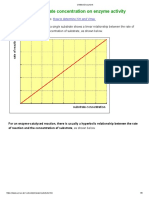
You might also like
- Enzyme KineticsDocument10 pagesEnzyme KineticsQuenneBelocuraNo ratings yet
- Michaelis Menten EquationDocument4 pagesMichaelis Menten Equationsumant5shringari100% (1)
- Enzyme Kinetics Study RatesDocument20 pagesEnzyme Kinetics Study RatesSirsha PattanayakNo ratings yet
- Michaelis Menten Equation SummaryDocument7 pagesMichaelis Menten Equation SummaryBüşra GültekinNo ratings yet
- Biochemistry NotesDocument14 pagesBiochemistry NotesJocabed RadillaNo ratings yet
- Enzyme KineticsDocument8 pagesEnzyme KineticsronojoysenguptaNo ratings yet
- LineweaverBurk PlotDocument17 pagesLineweaverBurk PlotJawadNo ratings yet
- Enzyme Terminology ExplainedDocument3 pagesEnzyme Terminology ExplainedcapricornchrissNo ratings yet
- Enzyme and Acid - Base CatalysisDocument64 pagesEnzyme and Acid - Base Catalysisbinseung skzNo ratings yet
- Lecture On EnzymesDocument48 pagesLecture On EnzymesMary Jean DiazNo ratings yet
- Enzyme M&LDocument6 pagesEnzyme M&LNethra Sasikumar100% (1)
- Chapter 7.7 Abbreviated Lecture NotesDocument27 pagesChapter 7.7 Abbreviated Lecture NotesZach MaxwellNo ratings yet
- Lecture 3 Notes - EnzymologyDocument9 pagesLecture 3 Notes - EnzymologyYi Gong100% (3)
- Enzyme InhibitionDocument4 pagesEnzyme InhibitionNisha ManasNo ratings yet
- Enzymes - Kinetics HyperbolaDocument12 pagesEnzymes - Kinetics HyperbolaprincesschemistNo ratings yet
- Biology 3601 Biochemistry Enzyme Kinetics Laboratory BackgroundDocument5 pagesBiology 3601 Biochemistry Enzyme Kinetics Laboratory Backgroundkgeorges27No ratings yet
- Enzyme Kinetics ExplainedDocument17 pagesEnzyme Kinetics Explainedsherif87No ratings yet
- Enzymes and Enzyme Kinetics - January 2021Document34 pagesEnzymes and Enzyme Kinetics - January 2021dhiraj more100% (1)
- Simple Kinetics of Enzyme ActionDocument7 pagesSimple Kinetics of Enzyme ActionRavindra Mani TiwariNo ratings yet
- Michaelis-Menten Type Kinetics: E S) ES) K K K E S) ES)Document2 pagesMichaelis-Menten Type Kinetics: E S) ES) K K K E S) ES)fintastellaNo ratings yet
- Biology Biomolecules Part 2Document14 pagesBiology Biomolecules Part 2Ralph Kenneth LunaNo ratings yet
- Lecture 15: Enzyme Inhibition: Biochemistry I Fall Term, 2003Document4 pagesLecture 15: Enzyme Inhibition: Biochemistry I Fall Term, 2003archerofthestarsNo ratings yet
- EnzymeKinetics by P.C. Misra Professor, Department of Biochemistry Lucknow University, Lucknow-226 007Document21 pagesEnzymeKinetics by P.C. Misra Professor, Department of Biochemistry Lucknow University, Lucknow-226 007Dr. SHIVA AITHALNo ratings yet
- IIPE Class 4 24.03Document41 pagesIIPE Class 4 24.03bharatNo ratings yet
- 10-11, Kinetics of Enzyme Catalyzed ReactionDocument34 pages10-11, Kinetics of Enzyme Catalyzed ReactionS. Ansary100% (1)
- Enzyme-Kinetics and Specificity: Modified by Chung-Ming Lin Department of Biotechnology Ming Chuan UniversityDocument58 pagesEnzyme-Kinetics and Specificity: Modified by Chung-Ming Lin Department of Biotechnology Ming Chuan University陳家琪No ratings yet
- BS 3Document60 pagesBS 3himanshu_agraNo ratings yet
- Derivations OF: Enzyme Kinetics (I)Document20 pagesDerivations OF: Enzyme Kinetics (I)Sabari Krishnan B B100% (1)
- Slide Biop Enz KinDocument19 pagesSlide Biop Enz KinFahmi Ramadhan PutraNo ratings yet
- Slide Biop 6-Enz KinDocument19 pagesSlide Biop 6-Enz KinAri FaisalNo ratings yet
- Enzyme NotesDocument18 pagesEnzyme NotesDBPNo ratings yet
- Enzyme Kinetics GuideDocument26 pagesEnzyme Kinetics GuideToga Brandon100% (1)
- Michaelis Menten DerivedDocument4 pagesMichaelis Menten DerivedGOOGLI TECNo ratings yet
- Enzymes: Properties and KineticsDocument59 pagesEnzymes: Properties and Kineticsthamizh555100% (2)
- Enzyme Part 1-5 Vikneswaran 260110132004Document7 pagesEnzyme Part 1-5 Vikneswaran 260110132004Vikneswaran VîçkýNo ratings yet
- Teknik ReaksiDocument21 pagesTeknik ReaksiAhmadi FachryNo ratings yet
- Types of Inhibirtion GreatDocument3 pagesTypes of Inhibirtion Greatteam TSOTARENo ratings yet
- Enzyme2 2012Document45 pagesEnzyme2 2012Surya Prakash KabiNo ratings yet
- Some Points On Briggs-Haldane Kinetics - Meaning of The ParametersDocument5 pagesSome Points On Briggs-Haldane Kinetics - Meaning of The ParametersmousumimamunNo ratings yet
- Biochem ConceptsDocument9 pagesBiochem Conceptsdanena88No ratings yet
- Determining Km and Vmax for Enzyme ActivityDocument8 pagesDetermining Km and Vmax for Enzyme ActivityMOSES MILLERNo ratings yet
- Enzyme Catalysis: Mechanisms and Factors that Accelerate Reaction RatesDocument14 pagesEnzyme Catalysis: Mechanisms and Factors that Accelerate Reaction RatesAVNo ratings yet
- Kinetics of EnzymesDocument2 pagesKinetics of EnzymesJohnNo ratings yet
- Non-Linear Pharmacokinetics: Biopharmaceutics (Ashik Sir)Document9 pagesNon-Linear Pharmacokinetics: Biopharmaceutics (Ashik Sir)Hasan Al MasudNo ratings yet
- Chapter8&9 Enzyme KineticsDocument68 pagesChapter8&9 Enzyme KineticsSyuhadah NoordinNo ratings yet
- Lecture 12 - Enzyme Kinetics IDocument30 pagesLecture 12 - Enzyme Kinetics IThomas Jones50% (2)
- Enzyme Catalysis PDFDocument17 pagesEnzyme Catalysis PDFuzzal ahmedNo ratings yet
- Bio Chapter 3Document103 pagesBio Chapter 3Areej AlissaNo ratings yet
- Enzymology (GEB-2101) Lecture 3Document15 pagesEnzymology (GEB-2101) Lecture 3Niloy GhoshNo ratings yet
- Derivation of Reaction Rate Expression With Equilibrium AssumptionDocument6 pagesDerivation of Reaction Rate Expression With Equilibrium AssumptionViaTS13No ratings yet
- Enzyme Kinetics and Catalysis IIDocument36 pagesEnzyme Kinetics and Catalysis IIlsueyinNo ratings yet
- Enzyme Kinetics ExplainedDocument43 pagesEnzyme Kinetics ExplainedBlessings Chawinga100% (1)
- AsaDocument35 pagesAsamarz95No ratings yet
- MRQ_#3 Enzyme Kinetics_2015Document44 pagesMRQ_#3 Enzyme Kinetics_2015yanuar setiawanNo ratings yet
- Dr. Nazir Ahmad: Nehal BilalDocument15 pagesDr. Nazir Ahmad: Nehal BilalAayat MughalNo ratings yet
- Biophysj00128 0003Document8 pagesBiophysj00128 0003Dean AstumianNo ratings yet
- Mechaelis-Menten KineticsDocument7 pagesMechaelis-Menten KineticsfayeNo ratings yet
- Allosteric Enzyme Regulation ExplainedDocument4 pagesAllosteric Enzyme Regulation ExplainedabassNo ratings yet
- Theory and Applications of the Empirical Valence Bond Approach: From Physical Chemistry to Chemical BiologyFrom EverandTheory and Applications of the Empirical Valence Bond Approach: From Physical Chemistry to Chemical BiologyFernanda DuarteNo ratings yet
- Reviews in Computational ChemistryFrom EverandReviews in Computational ChemistryAbby L. ParrillNo ratings yet
- Guideline Specifications Test Procedures Acceptance Criteria Herbal Substances Herbal Preparations enDocument25 pagesGuideline Specifications Test Procedures Acceptance Criteria Herbal Substances Herbal Preparations enazeempharmacist4773No ratings yet
- VLT VMT Pump Brochure enDocument4 pagesVLT VMT Pump Brochure enLuis YañezNo ratings yet
- Calciumlactate 1Document2 pagesCalciumlactate 1JilNo ratings yet
- Test Pack-D5bf-4637-A89d-2df0b6233e93-150828025754-Lva1-App6892Document37 pagesTest Pack-D5bf-4637-A89d-2df0b6233e93-150828025754-Lva1-App6892Ram L Rao100% (1)
- Chapter17 (Chemical Equilibria)Document61 pagesChapter17 (Chemical Equilibria)joshuaNo ratings yet
- Lucio WP4 - Weatherproof R1Document2 pagesLucio WP4 - Weatherproof R1Shameer MajidNo ratings yet
- R18 Jntu SyllabusDocument31 pagesR18 Jntu SyllabusSnehith SagarNo ratings yet
- Phosphoric Acid ManualDocument33 pagesPhosphoric Acid ManualAbi Ponce Mena86% (7)
- Manipal Institute of Technology: Subject: Engineering Physics (Phy101/102) Time: 3 Hrs. Max. Marks: 50Document6 pagesManipal Institute of Technology: Subject: Engineering Physics (Phy101/102) Time: 3 Hrs. Max. Marks: 50Chhayank AnkurNo ratings yet
- Chapter 2Document49 pagesChapter 2Umesh Chandra100% (1)
- Mil F 24787Document22 pagesMil F 24787ScottSherodNo ratings yet
- SGC - Commodities & HS Codes - AJCEPT VJEPA 2015 From April 2015Document2 pagesSGC - Commodities & HS Codes - AJCEPT VJEPA 2015 From April 2015Nguyen Thanh PhuongNo ratings yet
- Safe and Easy Water Release: Quick Transfer To KF TitratorsDocument2 pagesSafe and Easy Water Release: Quick Transfer To KF Titratorsmohammed younesNo ratings yet
- NVABR1014-Sealant SealantEquipment PDFDocument8 pagesNVABR1014-Sealant SealantEquipment PDFsterlingNo ratings yet
- Full Text ThesisDocument219 pagesFull Text ThesisVishal SalveNo ratings yet
- AIRAH Heat Load Fact SheetDocument3 pagesAIRAH Heat Load Fact Sheetvarshneyrk@rediffmail.comNo ratings yet
- Foam Sampling Testing ProcedureDocument1 pageFoam Sampling Testing ProcedurezahasherNo ratings yet
- PT. Sukses Mitra Silika Material Safety Data SheetDocument10 pagesPT. Sukses Mitra Silika Material Safety Data SheetRiko Adi SunaryoNo ratings yet
- Assignment For Chemical Kinetics: Submitted To: Dr. Erum JabeenDocument11 pagesAssignment For Chemical Kinetics: Submitted To: Dr. Erum JabeenIsrar KhanNo ratings yet
- Lab Report 2 Fruit BatteryDocument8 pagesLab Report 2 Fruit BatteryJullifer TubaNo ratings yet
- IS2386-Part3-1963 (Reaffirmed2021) - AGGREGATES FOR CONCRETE-SPECIFIC GRAVITY - DENSITY - VOIDS - ABSORPTION - BULKINGDocument22 pagesIS2386-Part3-1963 (Reaffirmed2021) - AGGREGATES FOR CONCRETE-SPECIFIC GRAVITY - DENSITY - VOIDS - ABSORPTION - BULKINGbaubleNo ratings yet
- 1.nano - Thin FilmDocument10 pages1.nano - Thin FilmTJPRC PublicationsNo ratings yet
- Sluice Valves - Pljosnati ZasuniDocument8 pagesSluice Valves - Pljosnati ZasuniKemal GokovicNo ratings yet
- Media FilterDocument1 pageMedia FilterahmadNo ratings yet
- A Review of Literature For The Structural Assessment of Mitred Bends PDFDocument20 pagesA Review of Literature For The Structural Assessment of Mitred Bends PDFaemis2010No ratings yet
- Is.10742.1983 Gun Metal Rods & BarsDocument13 pagesIs.10742.1983 Gun Metal Rods & Barsjra9090No ratings yet
- Casein PlasticDocument6 pagesCasein Plasticpiepie2No ratings yet
- How It Works: Double Block and Bleed and Double Isolation-And-Bleed ValvesDocument2 pagesHow It Works: Double Block and Bleed and Double Isolation-And-Bleed ValvesTiffany DacinoNo ratings yet