You might also like
- Efectos No Cardiovasculares Asociados A EstatinasDocument10 pagesEfectos No Cardiovasculares Asociados A EstatinasMacv MclNo ratings yet
- Rhabdomiolisis 2008 PDFDocument7 pagesRhabdomiolisis 2008 PDFMacv MclNo ratings yet
- 40 - Practical Approach To The Febrile Child in The Emergency DeptDocument11 pages40 - Practical Approach To The Febrile Child in The Emergency DeptMacv MclNo ratings yet
- Enf Coronaria y RabdomiolisisDocument6 pagesEnf Coronaria y RabdomiolisisMacv MclNo ratings yet
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5783)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (890)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (72)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Angiography 170411210116Document44 pagesAngiography 170411210116Ndikintum MaciasNo ratings yet
- Pulmonary Embolism: Ahmad Faeez Aleeffuddin Bin Mohamad Bidin Mentor:Encik SamsuddinDocument22 pagesPulmonary Embolism: Ahmad Faeez Aleeffuddin Bin Mohamad Bidin Mentor:Encik SamsuddinJuni Zuhairah Abd GhafarNo ratings yet
- Different Degradation Rates of Nanofiber Vascular Grafts in Small and Large AnimalDocument35 pagesDifferent Degradation Rates of Nanofiber Vascular Grafts in Small and Large AnimalMadalina CalcanNo ratings yet
- Sop Rabbit Blood Collection Marginal Ear VeinDocument5 pagesSop Rabbit Blood Collection Marginal Ear Veinbappy bayzidNo ratings yet
- Relationship Between CKD Stage and Pulmonary Edema on Chest X-RayDocument6 pagesRelationship Between CKD Stage and Pulmonary Edema on Chest X-RayAnnisa RabbaniNo ratings yet
- Đề số 11Document6 pagesĐề số 11Quynh Anh NguyenNo ratings yet
- Body SystemsDocument116 pagesBody SystemskarunaNo ratings yet
- Wbi11 01 Rms 20220818Document28 pagesWbi11 01 Rms 20220818Tamanna Adiba100% (1)
- Cardiovascular SystemDocument10 pagesCardiovascular SystemitsmesuvekshaNo ratings yet
- Cambridge Primary Science Teacher's Resource Book 6 With CD-ROM, Fiona Baxter and Liz Dilley, Cambridge University Press - PublicDocument38 pagesCambridge Primary Science Teacher's Resource Book 6 With CD-ROM, Fiona Baxter and Liz Dilley, Cambridge University Press - PublicSaleha Shoaib68% (22)
- CVSDocument58 pagesCVSDoina SvetNo ratings yet
- Nursing Care Plan Priorities for Hemodialysis PatientsDocument6 pagesNursing Care Plan Priorities for Hemodialysis PatientsPauling FrezNo ratings yet
- Addicted Heroine NovelDocument283 pagesAddicted Heroine NovelPanktiNo ratings yet
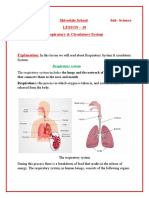
- Respiratory & Circulatory SystemDocument12 pagesRespiratory & Circulatory SystemYaSHVaRDHaN DaGaNo ratings yet
- Medical Terminology Cardiovascular SystemDocument35 pagesMedical Terminology Cardiovascular Systemapi-26819951486% (7)
- ATPL Human PerformanceDocument583 pagesATPL Human PerformanceAmilton Morillas100% (1)
- Anatomy and Physiology of DengueDocument4 pagesAnatomy and Physiology of Dengueciarkai_18No ratings yet
- Life Process: 1. Objective QuestionsDocument16 pagesLife Process: 1. Objective Questionsnavin gupta100% (1)
- Title Card Guide Card Activity Card Assessment Card Enrichment Card Tosumitup Reference Card Answer CardDocument19 pagesTitle Card Guide Card Activity Card Assessment Card Enrichment Card Tosumitup Reference Card Answer CardMyra Roa GasconNo ratings yet
- Anatomy of the Cardiovascular SystemDocument75 pagesAnatomy of the Cardiovascular SystemM RazzaqNo ratings yet
- Inotropes in Term NeonatesDocument6 pagesInotropes in Term Neonateserika lunaNo ratings yet
- Thrombosis and EmbolismDocument39 pagesThrombosis and Embolismilva100% (1)
- Dr. Naitik Trivedi & Dr. Upama Trivedi: Haemopoietic SystemDocument22 pagesDr. Naitik Trivedi & Dr. Upama Trivedi: Haemopoietic SystemthiruklpNo ratings yet
- MS300 Owners ManualDocument36 pagesMS300 Owners ManualAfdan FirdausNo ratings yet
- Practice Paper For MD1 2021 - AnswersDocument10 pagesPractice Paper For MD1 2021 - AnswersAlex TaylorNo ratings yet
- A Case Study On Ischemic Stroke: J.H. Cerilles State CollegeDocument21 pagesA Case Study On Ischemic Stroke: J.H. Cerilles State CollegeNicole Aranding100% (4)
- Dysrhythmia InterpretationDocument56 pagesDysrhythmia InterpretationgerajassoNo ratings yet
- PDA Journal SeptemberDocument19 pagesPDA Journal SeptemberZia100% (1)
- Bio Chapter 4Document5 pagesBio Chapter 4Aleeya MaisarahNo ratings yet
- 1 Cardiovascular SystemDocument51 pages1 Cardiovascular SystemChryzl JoyNo ratings yet