You might also like
- Ejercicios FQ3Document37 pagesEjercicios FQ3Instituto SosNo ratings yet
- Procedimiento soldadura y oxicorteDocument13 pagesProcedimiento soldadura y oxicorteLeonardo VenegasNo ratings yet
- AtmósferaDocument14 pagesAtmósferavivianaNo ratings yet
- Monografía - Producción de Bioetanol - XV Curso de TitulaciónDocument96 pagesMonografía - Producción de Bioetanol - XV Curso de TitulaciónRomulo Aycachi Inga100% (5)
- El ciclo hidrológico del aguaDocument2 pagesEl ciclo hidrológico del aguaJessica Ramírez MünzenmayerNo ratings yet
- Binomio de Newton - El Grado de Un Polinomio1Document3 pagesBinomio de Newton - El Grado de Un Polinomio1luisda_32No ratings yet
- ENCEFALO2Document69 pagesENCEFALO2luisda_32No ratings yet
- ENCEFALO1Document43 pagesENCEFALO1luisda_32No ratings yet
- EspermogramaDocument11 pagesEspermogramadragorexNo ratings yet
- Anteproyecto Ley Auge VIHSIDADocument33 pagesAnteproyecto Ley Auge VIHSIDAluisda_32No ratings yet
- Preguntas de preparación para examen de gastroenterología del IEESSDocument42 pagesPreguntas de preparación para examen de gastroenterología del IEESSCarlos Vinicio Erazo50% (2)
- Preguntas de preparación para examen de gastroenterología del IEESSDocument42 pagesPreguntas de preparación para examen de gastroenterología del IEESSCarlos Vinicio Erazo50% (2)
- Acidos NucleicosDocument9 pagesAcidos Nucleicosluisda_32No ratings yet
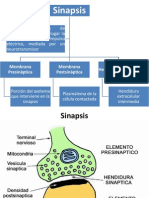
- Sin A Psis 2Document10 pagesSin A Psis 2luisda_32No ratings yet
- Ficha Técnica de SeguridadDocument2 pagesFicha Técnica de SeguridadYeraldin EspañaNo ratings yet
- Tarea N°3 MicrobiologiaDocument36 pagesTarea N°3 MicrobiologiaJasiel SolisNo ratings yet
- Practica #4 Punto de Fusion y EbullicionDocument7 pagesPractica #4 Punto de Fusion y EbullicionBryan Almanza Marcapura100% (1)
- Tabla de Alcanos Alquenos y AlquinosDocument4 pagesTabla de Alcanos Alquenos y AlquinosCch Quimica100% (5)
- Puntos de FusiónDocument8 pagesPuntos de FusiónAnonymous TqrFia7No ratings yet
- Tarea 1-Transferencia de Masa.Document33 pagesTarea 1-Transferencia de Masa.Hauslerr Mendoza LingánNo ratings yet
- Parches Piezoeléctricos Janus Impresos en 3D para La Eliminación de Bacterias Sonodinámicas y La Cicatrización de HeridasDocument20 pagesParches Piezoeléctricos Janus Impresos en 3D para La Eliminación de Bacterias Sonodinámicas y La Cicatrización de HeridasabiNo ratings yet
- Diapositivas de Carbohidratos (Bromatologia)Document15 pagesDiapositivas de Carbohidratos (Bromatologia)Jhonfredy Danielsmuños100% (1)
- IncandescenciaDocument5 pagesIncandescenciaalbertoacosNo ratings yet
- Obtención de BenzoinaDocument18 pagesObtención de BenzoinaDiana JulietaNo ratings yet
- ES Dynalife HTDocument2 pagesES Dynalife HTVictor ZhicayNo ratings yet
- ALMIDONDocument8 pagesALMIDONMaryzol CUevazz ZanchezzNo ratings yet
- Por qué se celebra el 16 de setiembre el Día Internacional de la Protección de la Capa de OzonoDocument2 pagesPor qué se celebra el 16 de setiembre el Día Internacional de la Protección de la Capa de OzonoBrus Condori CcoraNo ratings yet
- 2.3:2.4 Incendios de InteriorDocument66 pages2.3:2.4 Incendios de Interiorovillalba_25No ratings yet
- Informe de Trimetroprim Con SulfametoxazolDocument5 pagesInforme de Trimetroprim Con SulfametoxazolJerzy AmbrizNo ratings yet
- Oxidos de AzufreDocument19 pagesOxidos de AzufreJonathan Yesid Roa JaimesNo ratings yet
- GUÍA DBA 3 de Dónde Viene y Hacia Donde Fluye La Energía en Los Ecosistemas PDFDocument14 pagesGUÍA DBA 3 de Dónde Viene y Hacia Donde Fluye La Energía en Los Ecosistemas PDFSandra PachecoNo ratings yet
- Cinética de la reacción de nitrosación de la dimetil ureaDocument19 pagesCinética de la reacción de nitrosación de la dimetil ureaFrancisco LaraNo ratings yet
- Biologia 1Document27 pagesBiologia 1Aksel SpírituNo ratings yet
- Manual QuimicaDocument51 pagesManual QuimicaJohnny Alvarez100% (1)
- Dioxido de CarbonoDocument1 pageDioxido de CarbonoIsrael GarciaNo ratings yet
- La Materia y Sus EstadosDocument3 pagesLa Materia y Sus Estadosrolando macias cruzNo ratings yet
- Trabajo Practico Microbioogia de SuelosDocument4 pagesTrabajo Practico Microbioogia de SuelosJohana Tejada100% (1)
- Wuolah-Resumen 11 PotenciometríaDocument4 pagesWuolah-Resumen 11 PotenciometríaelaNo ratings yet
- Fatiga Alto Nro de CiclosDocument24 pagesFatiga Alto Nro de CiclosMartin Chimenti - ARO SANo ratings yet