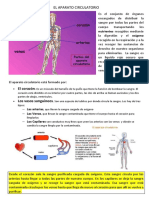
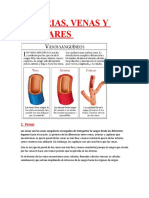
You might also like
- Discusion 4 Bioca IDocument22 pagesDiscusion 4 Bioca Iestradaf6No ratings yet
- Preguntas 6 y 8Document7 pagesPreguntas 6 y 8LIZETHNo ratings yet
- Acceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017From EverandAcceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017No ratings yet
- Radiofrecuencia en AliementosDocument16 pagesRadiofrecuencia en AliementosUlises Benjamin Narro Altamirano0% (1)
- Respuesta y Adaptación Fisiológica Al EjercicioDocument26 pagesRespuesta y Adaptación Fisiológica Al EjercicioSergio Valencia ZuñigaNo ratings yet
- Docente: Msc. Anabel González Siccha: Trujillo, 25 de Abril de 2014Document30 pagesDocente: Msc. Anabel González Siccha: Trujillo, 25 de Abril de 2014Shantall SaquicorayNo ratings yet
- T-9 Teoría Quimiosmótica. LanzaderasDocument24 pagesT-9 Teoría Quimiosmótica. LanzaderasAnonymous iIaMAfVNo ratings yet
- Eq.3 Fosforilacion Oxidativa 2Document24 pagesEq.3 Fosforilacion Oxidativa 2rubydonaldo0% (1)
- Dadores Aceptores y P/ODocument36 pagesDadores Aceptores y P/OFrancisco Abanto33% (3)
- Definición de Fosforilación OxidativaDocument10 pagesDefinición de Fosforilación OxidativaDiomary ReyesNo ratings yet
- Clase 17Document5 pagesClase 17Pedro Esteban Ovalle AburtoNo ratings yet
- Cadena Transportadora de Electrones y Fosforilacion OxidativaDocument29 pagesCadena Transportadora de Electrones y Fosforilacion OxidativaLoreto Carrasco AlarconNo ratings yet
- Monografia - Cadena Respiratoria y Fosforilación OxidativaDocument4 pagesMonografia - Cadena Respiratoria y Fosforilación OxidativaNoemi RadaNo ratings yet
- Fosforilacion Oxidativa Teoria QuimiosmoticaDocument18 pagesFosforilacion Oxidativa Teoria QuimiosmoticaJose Arceo CendejasNo ratings yet
- Bioquimica Practica - Cadena RespiratoriaDocument3 pagesBioquimica Practica - Cadena Respiratoriaingcivilunt1No ratings yet
- 3-2022 Cte y Fosforilacion OxidativaDocument56 pages3-2022 Cte y Fosforilacion OxidativaAbisai FuentesNo ratings yet
- Practica #4Document23 pagesPractica #4Eder Mariñas100% (1)
- Cadena Respiratoria y Fosforilación OxidativaDocument14 pagesCadena Respiratoria y Fosforilación OxidativaP. Arevalo IlianaNo ratings yet
- Fosforilación OxidativaDocument25 pagesFosforilación OxidativaEmilio Jeanpaul Mitacc GutierrezNo ratings yet
- Fosforilación OxidativaDocument20 pagesFosforilación OxidativaValeri Vasquez Cucho0% (1)
- Fosforilación Oxidativa - ResumenDocument10 pagesFosforilación Oxidativa - ResumenKevinDelaCruzNo ratings yet
- Fosforilación OxidativaDocument26 pagesFosforilación OxidativaXiomara Sanchez CahuanaNo ratings yet
- Discusion 4 Bioca IDocument21 pagesDiscusion 4 Bioca IElena Donado50% (4)
- Tema 8 y 9Document25 pagesTema 8 y 9halley moonNo ratings yet
- Mecanismo Quimio-Osmótico de La Producción de AtpDocument18 pagesMecanismo Quimio-Osmótico de La Producción de AtpKathe TafurNo ratings yet
- Transporte Electronico y Fosforilacion OxidativaDocument41 pagesTransporte Electronico y Fosforilacion OxidativanicolehurtadosNo ratings yet
- Universidad Catolica NordestanaDocument2 pagesUniversidad Catolica Nordestanacarlos magdiel abreu mendozaNo ratings yet
- Cuál Es La Estructura de La Cadena Respiratoria y Explique Su FuncionamientoDocument3 pagesCuál Es La Estructura de La Cadena Respiratoria y Explique Su FuncionamientoARIANA TAIS BRICE�O QUIROZNo ratings yet
- Tema 15 Cadena Respiratoria. Autor Alexander Sanchez Lamas.Document15 pagesTema 15 Cadena Respiratoria. Autor Alexander Sanchez Lamas.Alexander SanchezNo ratings yet
- Cadena de Trasporte y Fosforilacion KKDocument32 pagesCadena de Trasporte y Fosforilacion KKUriel AlvaradoNo ratings yet
- Citocromos y MitosisDocument14 pagesCitocromos y Mitosiswaldrapa29No ratings yet
- Nutrtición AnimalDocument14 pagesNutrtición AnimalYareli JimenezNo ratings yet
- Fosforilasa-Oxidativa-Corregido 2Document29 pagesFosforilasa-Oxidativa-Corregido 2Maksimum YgNo ratings yet
- Las MitocondriasDocument7 pagesLas MitocondriasMairaNo ratings yet
- Inhibidores y Desacopladores de La Cadena Respiratoria... 3º Ciclo ... AgronomíaDocument5 pagesInhibidores y Desacopladores de La Cadena Respiratoria... 3º Ciclo ... AgronomíaDarli Alvaro Camacho RojasNo ratings yet
- La Cadena Respiratoria y Fosforilación OxidativaDocument6 pagesLa Cadena Respiratoria y Fosforilación OxidativaCarlos VásquezNo ratings yet
- Fosforilacion OxidativaDocument27 pagesFosforilacion Oxidativajosuekstillo100% (4)
- Fosforilación OxidativaDocument2 pagesFosforilación OxidativaNEIVERNo ratings yet
- 06-Fosforilación OxidativaDocument4 pages06-Fosforilación OxidativaYasmin Morales LópezNo ratings yet
- 003 Bioenergética UC 2020Document30 pages003 Bioenergética UC 2020Helena QuintanaNo ratings yet
- Capítulo VIII - Conversión EnergéticaDocument4 pagesCapítulo VIII - Conversión EnergéticaFederico Rivadeneira CornejoNo ratings yet
- Informe Temario Bioquímica Ii 2021-2022 Primer ParcialDocument13 pagesInforme Temario Bioquímica Ii 2021-2022 Primer ParcialCinthya BenavidesNo ratings yet
- Cadena Respiración y Fosforilación Oxidativa Semana 8 CAE II 2022Document14 pagesCadena Respiración y Fosforilación Oxidativa Semana 8 CAE II 2022vimoniNo ratings yet
- MITOCONDDocument35 pagesMITOCONDLEYNER FERNANDEZ DIAZNo ratings yet
- 7.1 Fosforilacion Oxidativa y FotosforilacionDocument21 pages7.1 Fosforilacion Oxidativa y FotosforilacionDiana Diana ValenzuelaNo ratings yet
- CLASE 4 - Defi G6PD - DM - Cad Respiratoria DiabetesDocument38 pagesCLASE 4 - Defi G6PD - DM - Cad Respiratoria DiabetesOrions MTovalinoNo ratings yet
- Oxidacion BiologicaDocument42 pagesOxidacion Biologica10ivanleoNo ratings yet
- PresentaciónDocument21 pagesPresentaciónSugeiry FriasNo ratings yet
- Cadena RespiratoriaDocument27 pagesCadena RespiratoriaRicardo FonsecaNo ratings yet
- Unidad Ii. Cadena Respiratoria y Fosforilacion Oxidativa PDFDocument34 pagesUnidad Ii. Cadena Respiratoria y Fosforilacion Oxidativa PDFMafer AraujoNo ratings yet
- Control Respiratorio - Curvas de Consumo de OxígenoDocument12 pagesControl Respiratorio - Curvas de Consumo de OxígenoTahía TamaraNo ratings yet
- Inhibidores de La Cadena Transportadoras de ElectronesDocument13 pagesInhibidores de La Cadena Transportadoras de ElectronesRomina reyesNo ratings yet
- T7. Cadena Respiratoria-CLAVDocument53 pagesT7. Cadena Respiratoria-CLAVPaolo Gianfranco Luna Victoria GutierrezNo ratings yet
- Cadena Respiratoria Aporte Elctronico y Fosforilación OxidativaDocument40 pagesCadena Respiratoria Aporte Elctronico y Fosforilación Oxidativa悲しい エレン・イェーガーNo ratings yet
- Ashley Salcedo 100478923 Bioquimica 2Document5 pagesAshley Salcedo 100478923 Bioquimica 2Lenny MotaNo ratings yet
- Cadena Respiratoria y Fosforilación OxidativaDocument11 pagesCadena Respiratoria y Fosforilación OxidativaAldric CondoriNo ratings yet
- Fosforilación Oxidativa (Resumen)Document5 pagesFosforilación Oxidativa (Resumen)Alfonso2KNo ratings yet
- Cadena Respiratoria MitocondrialDocument7 pagesCadena Respiratoria Mitocondrialana guadalupe rodriguez ariasNo ratings yet
- Caso Clinico 2Document14 pagesCaso Clinico 2ANNIE NICOLL ZAVALETA QUIROZNo ratings yet
- Acceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012From EverandAcceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012No ratings yet
- Tipos de EscaldadoDocument4 pagesTipos de EscaldadoUlises Benjamin Narro Altamirano100% (1)
- Vias Glucolíticas Aerobia y AnaerobiaDocument17 pagesVias Glucolíticas Aerobia y AnaerobiaUlises Benjamin Narro AltamiranoNo ratings yet
- Tecnología Agroindustrial 1Document48 pagesTecnología Agroindustrial 1Ulises Benjamin Narro AltamiranoNo ratings yet
- CERVEZA DiapositivasDocument36 pagesCERVEZA DiapositivasUlises Benjamin Narro AltamiranoNo ratings yet
- Elaboracion de Mermelada de Fresa en La IndustriaDocument6 pagesElaboracion de Mermelada de Fresa en La IndustriaUlises Benjamin Narro Altamirano67% (3)
- Elaboracion de Durazno en AlmibarDocument8 pagesElaboracion de Durazno en AlmibarUlises Benjamin Narro AltamiranoNo ratings yet
- Clase 04 (02) - Catabolismo de ProteinaDocument11 pagesClase 04 (02) - Catabolismo de ProteinaUlises Benjamin Narro AltamiranoNo ratings yet
- Anatomia y Fisiologia Del CorazonDocument26 pagesAnatomia y Fisiologia Del CorazonJonathan Andres Franco SánchezNo ratings yet
- Fisiología Del Sistema Vascular 2017-I PDFDocument34 pagesFisiología Del Sistema Vascular 2017-I PDFRosa Lilian GarciaNo ratings yet
- 2 Metabolismo 1Document33 pages2 Metabolismo 1juan carlosNo ratings yet
- Eduardo Díaz Balderas Felipe López Estudillo Jehur Mastranzo Osorio Jayr Zamora RojasDocument27 pagesEduardo Díaz Balderas Felipe López Estudillo Jehur Mastranzo Osorio Jayr Zamora RojasJehur Mastranzo OsorioNo ratings yet
- MúsculosDocument20 pagesMúsculosJorge Enrique Hernández NaranjoNo ratings yet
- Instructivo N. °02-Accion de Inhibidores Enzimaticos en La Cadena RespiratoriaDocument8 pagesInstructivo N. °02-Accion de Inhibidores Enzimaticos en La Cadena RespiratoriaNataly ToribioNo ratings yet
- Reacciones de Desdoblamiento de La GlucosaDocument3 pagesReacciones de Desdoblamiento de La GlucosaRuben SucuzhañayNo ratings yet
- Cadena Respiratoria y Fosforilación OxidativaDocument18 pagesCadena Respiratoria y Fosforilación OxidativaSantimioNo ratings yet
- Sopa de LetrasDocument1 pageSopa de Letrascarlos andres ballen castañoNo ratings yet
- Actividad 1.16. Mapa Conceptual Sobre Respiración Celular AeróbicaDocument2 pagesActividad 1.16. Mapa Conceptual Sobre Respiración Celular AeróbicaObed EspinosaNo ratings yet
- Corazón PDFDocument11 pagesCorazón PDFpepeNo ratings yet
- Apuntes TEMA 8 - Metabolismo OxidativoDocument27 pagesApuntes TEMA 8 - Metabolismo OxidativoSiham LopezNo ratings yet
- Harper Examen Cadena Respiratoria y Fosforilacion OxidativaDocument3 pagesHarper Examen Cadena Respiratoria y Fosforilacion OxidativaSissi Gryzbowski Gainza100% (3)
- Metabolismo Central Glucolisis Gluconeogenesis Oxidacion Del Piruvato Metabolismo Del Glicogeno Ciclo de Krebs 97316 Downloable 1564396Document23 pagesMetabolismo Central Glucolisis Gluconeogenesis Oxidacion Del Piruvato Metabolismo Del Glicogeno Ciclo de Krebs 97316 Downloable 1564396Mariana TrejoNo ratings yet
- Guia 7 PDH, CK, CTE y FO Biol166 2020Document4 pagesGuia 7 PDH, CK, CTE y FO Biol166 2020claudia cartesNo ratings yet
- Informe Mitocondrias.Document19 pagesInforme Mitocondrias.Soledad ZapataNo ratings yet
- Bioenergetica y MetabolismoDocument40 pagesBioenergetica y Metabolismonct127No ratings yet
- Carbohidratos IIDocument71 pagesCarbohidratos IISara Mogrovejo AguirreNo ratings yet
- Recopilado UnalmDocument2 pagesRecopilado UnalmFelix IturrizagaNo ratings yet
- Sistema CardiovascularDocument41 pagesSistema CardiovascularPaola AndreaNo ratings yet
- Describe La Respiración AnaeróbicaDocument3 pagesDescribe La Respiración AnaeróbicaSteice Gabriela Lescano OlivaNo ratings yet
- Arteria MuscularDocument8 pagesArteria MuscularDannery HernándezNo ratings yet
- 2.el Aparato CirculatorioDocument3 pages2.el Aparato CirculatorioYinmar GarciaNo ratings yet
- CICLO de KREBS (Diapositivas)Document27 pagesCICLO de KREBS (Diapositivas)eduardito100% (2)
- 3.5 Gasto CardiacoDocument6 pages3.5 Gasto CardiacoBlueAbeNo ratings yet
- Taller Conceptos HemodinamicaDocument11 pagesTaller Conceptos HemodinamicadeyvidNo ratings yet
- (Respiración Celular y Tarea)Document3 pages(Respiración Celular y Tarea)Carlos Antonio Mendoza ArteagaNo ratings yet
- EL APARATO CIRCULATORIO 2º PrimariaDocument2 pagesEL APARATO CIRCULATORIO 2º PrimariaM Carmen Esteban MartínNo ratings yet
- Arteria Vena y CapilarDocument4 pagesArteria Vena y Capilarmarianita tumumNo ratings yet