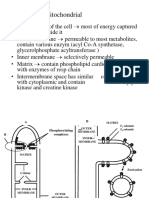
You might also like
- Mitochondrial oxidative phosphorylation and the proton gradientDocument47 pagesMitochondrial oxidative phosphorylation and the proton gradientSadaf BegNo ratings yet
- 13 OxphosDocument47 pages13 OxphosRahulNo ratings yet
- Oxidative Phosphorylation: Molecular Biochemistry IDocument47 pagesOxidative Phosphorylation: Molecular Biochemistry InithinkatzNo ratings yet
- Oxidative Phosphorylation: Molecular Biochemistry IDocument39 pagesOxidative Phosphorylation: Molecular Biochemistry IDozdiNo ratings yet
- Oxidative Phosphorylation and Electron Transport Chain II Respiratory Complexes Structure and Organization Chemiosmotic TheoryDocument20 pagesOxidative Phosphorylation and Electron Transport Chain II Respiratory Complexes Structure and Organization Chemiosmotic TheoryIffatnazNo ratings yet
- BotanyDocument2 pagesBotanyIrish Diana De AldayNo ratings yet
- Electron Transport System: Chemiosmotic Theory and Proton PumpingDocument58 pagesElectron Transport System: Chemiosmotic Theory and Proton PumpingSantosh KumarNo ratings yet
- 4 Bioenergetics and Oxidative Metabolism IIDocument3 pages4 Bioenergetics and Oxidative Metabolism IILinus LiuNo ratings yet
- Respiratory Chain & Oxidative Phosphorylation TutorialDocument57 pagesRespiratory Chain & Oxidative Phosphorylation TutorialHanifa AffianiNo ratings yet
- BBC2 K27,28RCDocument42 pagesBBC2 K27,28RCMarieta RitongaNo ratings yet
- Electron Transport and Oxidative PhosphorylationDocument34 pagesElectron Transport and Oxidative PhosphorylationAbeWatanabeNo ratings yet
- Proton Translocation Pathway: ROS Production (Reviewed in (2) )Document2 pagesProton Translocation Pathway: ROS Production (Reviewed in (2) )050Alya NurNo ratings yet
- Electron Transport and Oxidative Phosphorylation ExplainedDocument22 pagesElectron Transport and Oxidative Phosphorylation ExplainedAs ShahirahNo ratings yet
- Cellular Respiration Electron Transport ChainDocument24 pagesCellular Respiration Electron Transport ChainDark_KiroNo ratings yet
- Electron Transport ChainDocument33 pagesElectron Transport ChainAfaq AhmadNo ratings yet
- ChemiosmosisDocument7 pagesChemiosmosisSkenzKenzNo ratings yet
- Oxidative Phosphorylation 1Document22 pagesOxidative Phosphorylation 1fatin nadia100% (1)
- Photosystems I and II Light ReactionsDocument8 pagesPhotosystems I and II Light ReactionsTinna Devi ArmasamyNo ratings yet
- Electron Transport Chain ExplainedDocument9 pagesElectron Transport Chain Explainedmaria genioNo ratings yet
- Gayatree PaniDocument23 pagesGayatree PaniGobindaSahuNo ratings yet
- Electron Transport Chain - 1Document7 pagesElectron Transport Chain - 1Manash SarmahNo ratings yet
- Chapter 20 ETC and Oxidative PhosphDocument29 pagesChapter 20 ETC and Oxidative PhosphSpencer ThomasNo ratings yet
- Etc PDFDocument14 pagesEtc PDFjamalNo ratings yet
- Mitochondria: Mitochondria and Oxidative Phosphorylation Molecular Mechanisms of Electron Transport and Proton PumpingDocument54 pagesMitochondria: Mitochondria and Oxidative Phosphorylation Molecular Mechanisms of Electron Transport and Proton Pumpingnokate konkoorNo ratings yet
- ch.20 Imp. Note + PyqDocument18 pagesch.20 Imp. Note + Pyqجمال محمود السيايدةNo ratings yet
- Electron Transport Chain BSBT025F18 PDFDocument7 pagesElectron Transport Chain BSBT025F18 PDFZaki SyedNo ratings yet
- In Relation To Lsm2101 Lec 2Document11 pagesIn Relation To Lsm2101 Lec 2jojolim18No ratings yet
- Oxidation Glucose Coenzymes Glycolysis Citric Acid Cycle: Electron TransportDocument4 pagesOxidation Glucose Coenzymes Glycolysis Citric Acid Cycle: Electron TransportHabibur RahamanNo ratings yet
- Electron Transport ChainDocument18 pagesElectron Transport ChainArlyn Pasion BungbongaNo ratings yet
- The Electron Transport System Also Called The Electron Transport ChainDocument5 pagesThe Electron Transport System Also Called The Electron Transport ChainRe UpNo ratings yet
- BiochemDocument10 pagesBiochemHoàng LâmNo ratings yet
- BIOLOGICAL OXIDATION VIA ETCDocument22 pagesBIOLOGICAL OXIDATION VIA ETCbluegreenalga100% (2)
- Electron Transport ChainDocument4 pagesElectron Transport ChainBae SeujiNo ratings yet
- ETC transports electrons to power ATPDocument9 pagesETC transports electrons to power ATPalokesh1982100% (1)
- Electron Transport and Oxidative PhosphorylationDocument34 pagesElectron Transport and Oxidative Phosphorylationapi-321453350No ratings yet
- 1 Oxidative PhosphorylationDocument10 pages1 Oxidative PhosphorylationRoland ToroNo ratings yet
- ASSIGNMENT NO2 ChemDocument5 pagesASSIGNMENT NO2 ChemD AmanatNo ratings yet
- Cellular Energy TransactionsDocument3 pagesCellular Energy TransactionsDrAmit VermaNo ratings yet
- 16.2.3 Oxidative Phosphorylation: Cellular RespirationDocument47 pages16.2.3 Oxidative Phosphorylation: Cellular Respirationeryna sofeaNo ratings yet
- Aerobic Respiration: Chemiosmosis and Electron Transport ChainDocument17 pagesAerobic Respiration: Chemiosmosis and Electron Transport ChainAihk kenneth BaronaNo ratings yet
- Electron Transport ch14Document21 pagesElectron Transport ch14tania.delafuenteNo ratings yet
- Electron TransportDocument59 pagesElectron TransportEllexie JoycenolNo ratings yet
- Electron Transport Chain - WikipediaDocument53 pagesElectron Transport Chain - WikipediaLsaurusNo ratings yet
- Electron TransportDocument8 pagesElectron Transportเทพนิมิตร สมภักดีNo ratings yet
- Biological Oxidation (Electron Transport Chain - Chemiosmosis - Oxidative Phosphorylation - Uncouplers)Document22 pagesBiological Oxidation (Electron Transport Chain - Chemiosmosis - Oxidative Phosphorylation - Uncouplers)Krishia GalonaNo ratings yet
- BiologyDocument14 pagesBiologyHarsh SharmaNo ratings yet
- Electron Transport ChainDocument3 pagesElectron Transport ChainEmma MelNo ratings yet
- Lehninger Principles of Biochemistry PDFDocument37 pagesLehninger Principles of Biochemistry PDFmonitamiftahNo ratings yet
- Oxidative PhosphorylationDocument28 pagesOxidative PhosphorylationAnbuNo ratings yet
- Electron Transport ChainetcDocument19 pagesElectron Transport Chainetcpk kaleenaNo ratings yet
- J. Biol. Chem. 2001 Matsuno Yagi 19006 11Document7 pagesJ. Biol. Chem. 2001 Matsuno Yagi 19006 11brianmore10No ratings yet
- Cellular RespirationDocument55 pagesCellular RespirationFuad sabsebNo ratings yet
- Electron Transport and Oxidative Phosphorylation: Refer To: Lehninger Principles of Biochemistry (Chapter 19)Document43 pagesElectron Transport and Oxidative Phosphorylation: Refer To: Lehninger Principles of Biochemistry (Chapter 19)Yousef KhallafNo ratings yet
- Quinone and Non-Quinone Redox Couples in Complex III: # Springer Science + Business Media, LLC 2008Document7 pagesQuinone and Non-Quinone Redox Couples in Complex III: # Springer Science + Business Media, LLC 2008abossyNo ratings yet
- Electrontransportchain 151027073626 Lva1 App6892Document29 pagesElectrontransportchain 151027073626 Lva1 App6892GobindaSahuNo ratings yet
- BIO 361 Exam 4 ReviewDocument45 pagesBIO 361 Exam 4 ReviewNigel Zhang100% (1)
- BIOCHEMICAL ENERGY PRODUCTION SAS PT 3Document5 pagesBIOCHEMICAL ENERGY PRODUCTION SAS PT 3Abegail Ashley PenoniaNo ratings yet
- Selected Constants: Oxidation–Reduction Potentials of Inorganic Substances in Aqueous SolutionFrom EverandSelected Constants: Oxidation–Reduction Potentials of Inorganic Substances in Aqueous SolutionNo ratings yet
- Nikon: Instruction ManualDocument58 pagesNikon: Instruction Manualabc007200No ratings yet
- Diabetes Mellitus Type I Insulin Secretions and Genetic DefectsDocument2 pagesDiabetes Mellitus Type I Insulin Secretions and Genetic Defectsabc007200No ratings yet
- ImmunosuppressionDocument1 pageImmunosuppressionabc007200No ratings yet
- Blood Coagulation System Overview & Key FactorsDocument4 pagesBlood Coagulation System Overview & Key Factorsabc007200100% (1)
- Cytotoxic Drugs in The Treatment of CancerDocument2 pagesCytotoxic Drugs in The Treatment of Cancerabc007200No ratings yet
- Yashica FX 3 FX 7Document16 pagesYashica FX 3 FX 7Amrina RosadaNo ratings yet
- Work of BreathingDocument32 pagesWork of Breathingabc007200No ratings yet
- Basic Pascal Syntax: in This ChapterDocument54 pagesBasic Pascal Syntax: in This ChapterEldar ČokićNo ratings yet
- Bcr-Abl: Cancer Protein Structure and Function: About This WorksheetDocument2 pagesBcr-Abl: Cancer Protein Structure and Function: About This WorksheetRed FoxNo ratings yet
- Chronic Hepatitis B NEJM 2022Document15 pagesChronic Hepatitis B NEJM 2022Πάνος ΣταγήςNo ratings yet
- Neuroscience For Kids - The SynapseDocument3 pagesNeuroscience For Kids - The SynapseChantel AceveroNo ratings yet
- Protein Nutrition (David Bender)Document68 pagesProtein Nutrition (David Bender)Anonymous 8h8Rw6YmAnNo ratings yet
- Genetics and InheritanceDocument33 pagesGenetics and InheritanceLawrence Onthuga100% (1)
- Ch15 Test File-Gene Mutation and Molecular MedicineDocument35 pagesCh15 Test File-Gene Mutation and Molecular MedicineDollar'sCornerNo ratings yet
- Science of Golden-Yeast Teacher-InstructionsDocument18 pagesScience of Golden-Yeast Teacher-InstructionsImam WicaksonoNo ratings yet
- The Pathophysiology of Intrahepatic Cholestasis of PregnancyDocument13 pagesThe Pathophysiology of Intrahepatic Cholestasis of PregnancyEnrique RosasNo ratings yet
- Physical Science SHS 8.1 ProteinsDocument21 pagesPhysical Science SHS 8.1 ProteinsjouselleduayNo ratings yet
- Wolpert, Lewis - How We Live and Why We DieDocument4 pagesWolpert, Lewis - How We Live and Why We DieThamires Ribeiro de MattosNo ratings yet
- Nucleotides, Nucleic Acids, and Heredity: Bettelheim / Brown / Campbell / Farrell / TorresDocument55 pagesNucleotides, Nucleic Acids, and Heredity: Bettelheim / Brown / Campbell / Farrell / TorresMica BernardoNo ratings yet
- Metabolism Food Label LabDocument5 pagesMetabolism Food Label LabEdda Jean Gumban Varona33% (3)
- Mce Igcse Biology PPT c18Document37 pagesMce Igcse Biology PPT c18elaineNo ratings yet
- MS MLPA Protocol One Tube MSP v008Document19 pagesMS MLPA Protocol One Tube MSP v008Sp PpvNo ratings yet
- Human Genome ProjectDocument33 pagesHuman Genome Projectsharmamaddy32No ratings yet
- Paul 2010Document18 pagesPaul 2010DianaMarcelaBurbanoNo ratings yet
- Transformation of E.coli With pGAL. Lab ReportqqDocument8 pagesTransformation of E.coli With pGAL. Lab ReportqqJawad Salehi0% (1)
- Complete Amino Acid Sequence of Human Serum AlbuminDocument4 pagesComplete Amino Acid Sequence of Human Serum AlbuminJustine May Balinggan DelmasNo ratings yet
- Binary and Co Integrative VectorsDocument25 pagesBinary and Co Integrative VectorsSakshi BuchkeNo ratings yet
- Bio 212 HW 1 KeyDocument6 pagesBio 212 HW 1 KeyMitter JonesNo ratings yet
- Module 2 - Genetic Engineering 1Document20 pagesModule 2 - Genetic Engineering 1Ash MiNo ratings yet
- Final PaperDocument6 pagesFinal PaperALOHRA COLLIN FEBRERONo ratings yet
- Sars Covid - Molecular Biology CT Value Result Test DescriptionDocument1 pageSars Covid - Molecular Biology CT Value Result Test DescriptionRam JaneNo ratings yet
- Practice Prelim 3Document9 pagesPractice Prelim 3EricaNo ratings yet
- Module 9B - Transcription and Post-Transcriptional ModificationDocument46 pagesModule 9B - Transcription and Post-Transcriptional Modificationgleni robredoNo ratings yet
- Bio01 Co2 PPT - Cell CycleDocument120 pagesBio01 Co2 PPT - Cell CycleMarc Ronald de LeonNo ratings yet
- Biology: Chromosomes, Genes and DNADocument38 pagesBiology: Chromosomes, Genes and DNAMaane BrunoNo ratings yet
- Quiz Medical GeneticsDocument43 pagesQuiz Medical GeneticsMedShare95% (19)
- Biotechnology - Igcse BiologyDocument4 pagesBiotechnology - Igcse BiologyTheStarNo ratings yet
- Looking Beyond The Monoamine Hypothesis: Johan A Den BoerDocument9 pagesLooking Beyond The Monoamine Hypothesis: Johan A Den BoerdonkeyendutNo ratings yet